Grupo parafilético de cetáceos primitivos desde el Eoceno temprano hasta el Oligoceno tardío.
Archaeoceti ("ballenas antiguas"), o Zeuglodontes en la literatura más antigua, es un grupo parafilético de cetáceos primitivos que vivieron desde el Eoceno temprano hasta el Oligoceno tardío ( hace 50 a 23 millones de años ). [1] Representan la radiación de cetáceos más antigua , incluyen las etapas anfibias iniciales en la evolución de los cetáceos , por lo que son los antepasados de los dos subórdenes de cetáceos modernos, Mysticeti y Odontoceti . [2] Esta diversificación inicial se produjo en las aguas poco profundas que separaron India y Asia hace 53 a 45 millones de años , lo que dio como resultado unas 30 especies adaptadas a una vida plenamente oceánica. La ecolocalización y la alimentación por filtración evolucionaron durante una segunda radiación hace 36 a 35 millones de años . [3]
Todos los arqueocetos del Ypresiense (56 a 47,8 millones de años) y la mayoría del Lutetiense (47,8 a 41,3 millones de años) se conocen exclusivamente en Indo-Pakistán, pero los géneros Bartoniano (41,3 a 38,0 millones de años) y Priaboniano (38,0 a 33,9 millones de años) se conocen en en toda la Tierra, incluyendo América del Norte, Egipto, Nueva Zelanda y Europa. Aunque no existe consenso sobre el modo de locomoción del que eran capaces los cetáceos durante el Luteciano tardío, era muy poco probable que estuvieran tan bien adaptados al océano abierto como los cetáceos vivos. Probablemente llegaron hasta América del Norte a lo largo de aguas costeras, ya sea alrededor de África y hasta América del Sur, o más probablemente, sobre el mar de Tetis (entre Eurasia y África) y a lo largo de las costas de Europa, Groenlandia y América del Norte. [4]
Pakicetus , un pakicetido (dibujo que muestra restos fósiles conservados).
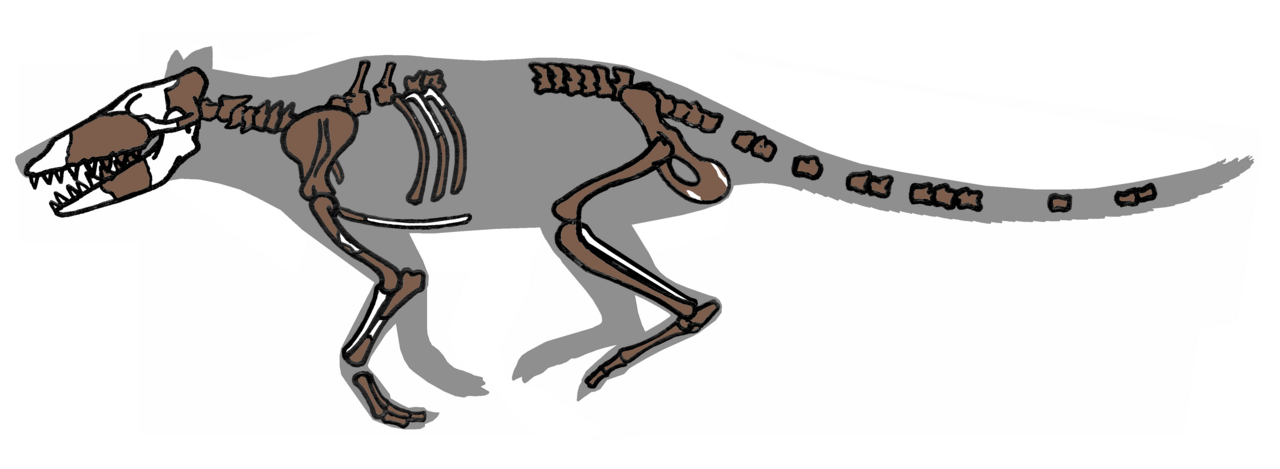
Identificados por primera vez como cetáceos por West 1980, los pakicetidos , la más arcaica de las ballenas, tenían patas largas y delgadas y una cola larga y estrecha, y podían alcanzar el tamaño de un lobo moderno. Sólo se han encontrado en sedimentos de corrientes de agua dulce en el noroeste de la India y el norte de Pakistán, y probablemente eran más limícolas que nadadores. [7]
Se conocen decenas de fósiles, pero sólo de cráneos, dientes y fragmentos de mandíbulas; no se han encontrado esqueletos completos. La dentición varió; las especies más pequeñas tenían dientes como los modernos comedores de pescado, y las más grandes se parecían más a las hienas modernas. Los pakicetidos pueden haber sido depredadores o alimentadores de carroña. Ni el cráneo ni la dentición de los pakicetidos se parecen a los de las ballenas modernas, pero el proceso sigmoideo, el involucro , los huesecillos paquiostóticos (compactos) y rotados de sus orejas aún revelan su naturaleza cetácea. [2]
Ambulocetidae
Ambulocetus , un ambulocétido (dibujo que muestra restos fósiles conservados).
La siguiente familia divergente de ballenas, los Ambulocetidae , eran grandes, ya totalmente acuáticas [8] y parecidas a cocodrilos, con patas grandes y una cola fuerte. Los sedimentos indican que vivían en zonas costeras y sus huesos compactos sugieren que eran depredadores de emboscada más que de persecución rápida. También conocidos exclusivamente de Pakistán e India, los ambulocétidos incluyen la ballena más antigua conocida, Himalayacetus , que se cree que tiene 53,5 millones de años , unos 4 millones de años más que el resto de su familia. [7]
De los menos de 10 fósiles que se han descrito, uno, Ambulocetus natans , está casi completo y es la principal fuente de información sobre la evolución temprana de los cetáceos. Del tamaño de un león marino macho, tenía una cabeza grande con un hocico largo y dientes robustos y muy desgastados. La mandíbula inferior muestra que Ambulocetus tenía un tejido blando inusual que conectaba la parte posterior de la mandíbula con el oído medio, un pequeño equivalente a la gran almohadilla grasa receptora de sonido de los odontocetos modernos. Sus ojos estaban colocados dorsalmente en la cabeza, pero miraban lateralmente. La musculatura de la cabeza, el cuello y la espalda era fuerte y la cola, sin aletas, era larga. Las extremidades traseras eran cortas, pero estaban provistas de patas largas. Las extremidades anteriores también eran cortas y estaban equipadas con cinco pezuñas cortas. Ambulocetus probablemente nadaba con sus patas traseras como una nutria moderna y era incapaz de soportar su propio peso en tierra. Probablemente era un cazador de emboscadas como los cocodrilos modernos. [2]
Remingtonocétidae
Kutchicetus , un remingtonocétido.
Los Remingtonocetidae tenían extremidades cortas y una cola fuerte y poderosa con vértebras aplanadas. Su largo hocico, sus ojos diminutos y la morfología de sus orejas sugieren que su visión era pobre y que el oído era su sentido dominante. También se han encontrado sólo en Pakistán y la India, y los sedimentos sugieren que vivían en aguas turbias en zonas costeras. Aunque probablemente podían vivir en tierra, aparentemente usaban sus colas para nadar. [7]
Se han descrito decenas de fósiles, pero la mayoría son sólo cráneos y mandíbulas inferiores con pocos restos dentales y poscraneales. Los remingtonocétidos probablemente variaron en tamaño y las especies más pequeñas coincidieron con Pakicetus y el Ambulocetus más grande . Los remingtonocétidos tenían hocicos más largos que otros arqueocetos, excepto que la morfología craneal también variaba considerablemente, probablemente reflejando diferentes dietas. Los ojos eran pequeños, pero las orejas grandes y muy separadas, probablemente reflejando un mayor énfasis en la audición bajo el agua. Los restos fragmentarios de poscráneos de remingtonocétidos sugieren que tenían un cuello largo y grandes extremidades traseras que probablemente podían soportar el peso del cuerpo en tierra. [2]
Las familias restantes y los cetáceos de la corona posteriores forman un clado unido por seis sinapomorfias : [9] El margen anterior de las fosas nasales externas se encuentra encima o detrás del tercer incisivo superior, la tribuna es ancha, los procesos supraorbitarios están presentes pero son cortos, el borde anterior de la órbita se encuentra por encima del segundo o tercer molar superior, la apófisis postorbitaria forma un ángulo de 90° con la cresta sagital y las vértebras cervicales son cortas.
Protocétidos
Maiacetus , un protocétido.
Los Protocetidae , conocidos tanto de África como de América, eran una familia diversificada con extremidades traseras y una cola fuerte, lo que indica que eran fuertes nadadores que colonizaron océanos poco profundos y cálidos, como los arrecifes. Afectaron en gran medida a la evolución de los cetáceos hace 47 a 41 millones de años , porque se extendieron por los océanos de la Tierra. [7] Tenían hocicos largos, ojos grandes y una abertura nasal ubicada más arriba en la cabeza que en los arqueocetos anteriores, lo que sugiere que podían respirar con la cabeza mantenida horizontalmente, similar a los cetáceos modernos, un primer paso hacia un espiráculo. Su dentición varió, pero comenzó a evolucionar hacia los dientes no masticadores de los cetáceos modernos, y probablemente eran cazadores activos. Su capacidad para moverse en tierra parece haber sido variable: en Rodhocetus y Peregocetus poseen una articulación sacroilíaca , lo que indica que podían moverse en tierra. [10] En otros géneros ( Georgiacetus y Aegicetus ), la pelvis no estaba conectada a la columna vertebral, lo que sugiere que las extremidades traseras no podrían haber soportado el peso corporal. [2] Algunos géneros ( Rohocetus ) tenían grandes patas traseras que formaban grandes remos, mientras que Aegicetus parece haber dependido más de su cola para impulsarse a través del agua. [11]
Basilosauridae
Dorudon , un basilosáurido.
Los basilosáuridos , que tenían diminutas extremidades traseras y extremidades delanteras en forma de aletas, eran obligatoriamente acuáticos y llegaron a dominar los océanos. Todavía carecían de la ecolocalización y las barbas de los odontocetos y misticetos modernos. [7] Los basilosáuridos y los dorudontidos son los cetáceos acuáticos obligados más antiguos de los que se conoce el esqueleto completo. Muestran una serie de adaptaciones acuáticas que no estaban presentes en los arqueocetos anteriores: [12] En la columna vertebral, las vértebras del cuello son cortas, las vértebras torácicas y lumbares tienen una longitud similar, las vértebras sacras no están fusionadas, las articulaciones sacroilíacas están ausentes y la cola corta tiene una vértebra esférica (lo que indica la presencia de una aleta). Las escápulas son anchas y en forma de abanico con acromiones anteriores y pequeñas fosas supraespinosas. Los cúbitos son grandes y tienen olécranons transversalmente planos, las muñecas y los antebrazos distales están aplanados en el plano de las manos y las extremidades traseras son pequeñas.
Taxonomía
Los Archaeoceti incluyen cinco familias bien establecidas : [13] El estatus de Kekenodontidae todavía está en disputa, y la familia se ubica en Archaeoceti, Mysticeti, [14] o incluso Delphinoidea. [15]
^ "Arqueoceti". Base de datos de paleobiología . Consultado el 20 de septiembre de 2018 .
^ abcde Thewissen 2002, págs. 36–8
^ Fordyce 2002, pag. 216
^ Geisler, Sanders y Luo 2005, Biogeografía, págs. 50-2
^ Steeman y col. 2009, pág. 573
^ Fordyce 2008, pag. 758
^ abcde Bajpai, Thewissen y Sahni 2009, pág. 675
^ Ando, Konami (2016). "Adiós a la vida terrestre - la fuerza torácica como nuevo indicador para determinar la paleoecología en mamíferos acuáticos secundarios". Revista de Anatomía . 229 (6): 768–777. doi :10.1111/joa.12518. PMC 5108153 . PMID 27396988.
^ Geisler, Sanders y Luo 2005, Discusión, p. 45
^ Olivier L, Bianucci G, Salas-Gismondi R, Di Celma C, Steurbaut E, Urbina M & de Muizon C (2019). "Una ballena anfibia del Eoceno Medio del Perú revela la dispersión temprana de cetáceos cuadrúpedos en el Pacífico Sur". Biología actual 29 (8): pág. 1352–1359.e3.
^ Gingerich PD, Antar HSH y Zalmout IS (2019). " Aegicetus gehennae , un nuevo protocétido del Eoceno tardío (Cetacea, Archaeoceti) de Wadi Al Hitan, Egipto, y la transición a la natación de ballenas con la cola". MÁS UNO 15 (3): e0230596
^ Thewissen y col. 2001, pág. 277
^ Rosa 2006, pag. 273
^ Clementz y col. 2014, figura 1
^ Gingerich 2005, tabla 15.1
Referencias
Andrews, CW (1919). "Una descripción de nuevas especies de zeuglodont y de tortuga coriácea del Eoceno del sur de Nigeria" (PDF) . Actas de la Sociedad Zoológica de Londres . 18 (3–4): 309–19. doi :10.1111/j.1096-3642.1919.tb02124.x. OCLC 228169273. Archivado desde el original (PDF) el 4 de enero de 2015 . Consultado el 1 de abril de 2013 .
Bajpai, Sunil; Gingerich, Philip D (diciembre de 1998). "Un nuevo arqueoceto del Eoceno (Mammalia, Cetacea) de la India y la época de origen de las ballenas". PNAS . 95 (26): 15464–68. Código bibliográfico : 1998PNAS...9515464B. doi : 10.1073/pnas.95.26.15464 . OCLC 678707438. PMC 28065 . PMID 9860991.
Bajpai, Sunil; Thewissen, JGM (2000). "Una nueva y diminuta ballena del Eoceno de Kachchh (Gujarat, India) y sus implicaciones para la evolución locomotora" (PDF) . Ciencia actual . 79 (10): 1478–82 . Consultado el 1 de julio de 2013 .
Bajpai, S; Thewissen, JG ; Sahni, A (2009). "El origen y evolución temprana de las ballenas: macroevolución documentada en el subcontinente indio" (PDF) . J Biosci . 34 (5): 673–86. doi :10.1007/s12038-009-0060-0. OCLC 565869881. PMID 20009264. S2CID 28232300 . Consultado el 1 de febrero de 2013 .
Bianucci, Giovanni; Gingerich, Philip D. (2011). " Aegyptocetus tarfa , n. gen. et sp. (Mammalia, Cetacea), del Eoceno medio de Egipto: clinorrinquia, olfato y audición en una ballena protocétida". Revista de Paleontología de Vertebrados . 31 (6): 1173–88. doi :10.1080/02724634.2011.607985. S2CID 85995809.
Clementz, Mark T.; Fordyce, R. Ewan; Mirar, Stephanie L.; Zorro, David L. (2014). "Antiguos isopaisajes marinos y evidencia isotópica de alimentación masiva por cetáceos del Oligoceno". Paleogeografía, Paleoclimatología, Paleoecología . 400 : 28–40. Código Bib : 2014PPP...400...28C. doi :10.1016/j.palaeo.2012.09.009.
Dehm, Richard; Oettingen-Spielberg, Therese zu (1958). Paläontologische und geologische Untersuchungen im Tertiär von Pakistan. 2. Die mitteleocänen Säugetiere von Ganda Kas bei Basal en el noroeste de Pakistán . Abhandlungen / Neue Folge, 91. Múnich: Beck. OCLC 163296508.
Flor, William Henry (1883). "Sobre la disposición de las órdenes y familias de mamíferos existentes". Actas de la Sociedad Zoológica de Londres : 178–86. OCLC 83091701.
Fordyce, Ewan (2002). "Evolución de los cetáceos". En Perrin, William R; Wiirsig, Bernd; Thewissen , JGM (eds.). Enciclopedia de mamíferos marinos . Prensa académica. págs. 214-25. ISBN 978-0-12-551340-1.
Fordyce, RE (2008). "Neoceti". En Perrin, WF; Wursig, B.; Thewissen , JGM (eds.). Enciclopedia de mamíferos marinos (2 ed.). Prensa académica. págs. 758–763. ISBN 978-0-12-373553-9.
Fraas, Eberhard (1904). "Neue Zeuglodonten aus dem unteren Mitteleocän vom Mokattam bei Cairo". Geologische und Paläontologische Abhandlungen . 6 (3): 199–220 . Consultado el 1 de julio de 2013 .
Geisler, Jonathan H; Lijadoras, Albert E; Luo, Zhe-Xi (julio de 2005). "Una nueva ballena protocétida (Cetacea, Archaeoceti) del Eoceno medio tardío de Carolina del Sur". Novitates del Museo Americano (3480): 1–68. doi :10.1206/0003-0082(2005)480[0001:ANPWCA]2.0.CO;2. hdl :2246/5671. OCLC 4630500767. S2CID 54060094.
Gervais, Paul (1876). "Comentarios sobre el tema del género Phocodon d'Agasiz" (PDF) . Revista de Zoología . 5 : 64–70 . Consultado el 1 de julio de 2013 .
Gibbes, Robert Wilson (1845). "Descripción de los dientes de un nuevo animal fósil encontrado en Green Sand de Carolina del Sur". Actas de la Academia de Ciencias Naturales de Filadelfia . 2 (9): 254–256 . Consultado el 1 de julio de 2013 .
"RWGibbes 1845". Paleobio DB .
Gingerich, PD (1992). "Mamíferos marinos (cetáceos y sirenia) del Eoceno de Gebel Mokattam y Fayum, Egipto: estratigrafía, edad y paleoambientes". Artículos sobre paleontología de la Universidad de Michigan . 30 : 1–84. hdl :2027.42/48630. OCLC 26941847.
Gingerich, Philip D. (2005). «Cetáceas» (PDF) . En Rose, KD; Archibald, JD (eds.). Mamíferos placentarios: origen, momento y relaciones de los principales clados existentes . Baltimore: Prensa de la Universidad Johns Hopkins. págs. 234-252 . Consultado el 1 de diciembre de 2013 .
Gingerich, Philip D (2007). "Stromerius nidensis, nuevo arqueoceto (Mammalia, Cetacea) de la Formación Qasr El-Sagha del Eoceno Superior, Fayum, Egipto" (PDF) . Aportaciones del Museo de Paleontología . 31 (13): 363–78. OCLC 214233870.
Gingerich, Philip D.; Arif, M.; Bhatti, M. Akram; Anwar, M.; Lijadoras, William J. (1997). " Basilosaurus drazindai y Basiloterus hussaini , nuevos Archaeoceti (Mammalia, Cetacea) de la Formación Drazinda del Eoceno Medio, con una interpretación revisada de las edades de los estratos balleneros en el grupo Kirthar de la cordillera Sulaiman, Punjab (Pakistán)". Contribuciones del Museo de Paleontología de la Universidad de Michigan . 30 (2): 55–81. hdl :2027.42/48652. OCLC 742731913.
Gingerich, Philip D.; Arif, Mahoma; Clyde, William C. (1995). "Nuevos Archaeocetes (Mammalia, Cetacea) de la Formación Domanda del Eoceno medio de la Cordillera Sulaiman, Punjab (Pakistán)". Contribuciones del Museo de Paleontología de la Universidad de Michigan . 29 (11): 291–330. hdl :2027.42/48650. OCLC 34123868.
Gingerich, Philip D.; Cappetta, Henri (2014). "Un nuevo arqueoceto y otros mamíferos marinos (Cetacea y Sirenia) de depósitos de fosfato del Eoceno medio inferior de Togo". Revista de Paleontología . 88 (1): 109-129. doi :10.1666/13-040. S2CID 85915213.
Gingerich, Philip D.; Raza, SM; Arif, M.; Anwar, M.; Zhou, X. (1994). "Nueva ballena del Eoceno de Pakistán y el origen de la natación de los cetáceos" (PDF) . Naturaleza . 368 (6474): 844–47. Código Bib :1994Natur.368..844G. doi :10.1038/368844a0. hdl : 2027.42/62571 . OCLC 742745707. S2CID 4315623.
Gingerich, Philip D.; Russell, Donald E. (1981). "Pakicetus inachus, un nuevo arqueoceto (Mammalia, Cetacea) de la formación Kuldana del Eoceno temprano-medio de Kohat (Pakistán)" (PDF) . Contribuciones del Museo de Paleontología, Museo de Michigan . 25 (11). OCLC 742729300 . Consultado el 20 de septiembre de 2018 .
Gingerich, Philip D.; Uhen, Mark D. (1996). "Ancalecetus simonsi, un nuevo arqueoceto dorudontino (Mammalia, Cetacea) del Eoceno tardío temprano de Wadi Hitan, Egipto" (PDF) . Contribuciones del Museo de Paleontología de la Universidad de Michigan . 29 (13): 359–401. OCLC 742731018 . Consultado el 1 de julio de 2013 .
Gingerich, Philip D; Ul-Haq, Munir; Khan, Intizar Hussain; Zalmout, Iyad S. (2001). "Estratigrafía del Eoceno y arqueocetos de ballenas (Mammalia, Cetacea) de Drug Lahar en la cordillera oriental de Sulaiman, Baluchistán (Pakistán)". Contribuciones del Museo de Paleontología de la Universidad de Michigan . 30 (11): 269–319. hdl :2027.42/48661. OCLC 50061585.
Gingerich, Philip D.; ul-Haq, Munir; von Koenigswald, Wighart; Sanders, William J.; Smith, B. Acebo; Zalmout, Iyad S. (2009). "Nueva ballena protocétida del Eoceno medio de Pakistán: nacimiento en tierra, desarrollo precocial y dimorfismo sexual". MÁS UNO . 4 (2): e4366. Código Bib : 2009PLoSO...4.4366G. doi : 10.1371/journal.pone.0004366 . OCLC 678622523. PMC 2629576 . PMID 19194487.
Gingerich, Philip D.; Ul-Haq, Munir; Zalmout, Iyad S.; Khan, Intizar Hussain; Malkani, S. (septiembre de 2001). "Origen de las ballenas de los primeros artiodáctilos: manos y pies de Protocetidae del Eoceno de Pakistán". Ciencia . 293 (5538): 2239–2242. Código Bib : 2001 Ciencia... 293.2239G. doi : 10.1126/ciencia.1063902 . ISSN 0036-8075. PMID 11567134. S2CID 21441797.
Gingerich, Philip D.; Zalmout, Iyad S.; Ul-Haq, Munir; Bhatti, M. Akram (2005). "Makaracetus bidens, un nuevo arqueoceto protocétido (Mammalia, Cetacea) del Eoceno medio temprano de Baluchistán (Pakistán)" (PDF) . Aportaciones del Museo de Paleontología . 31 (9): 197–210. OCLC 742723177 . Consultado el 1 de marzo de 2013 .
Goldin, Pavel; Zvonok, Evgenij (2013). " Basilotritus uheni , un nuevo cetáceo (Cetacea, Basilosauridae) del Eoceno medio tardío de Europa del Este". Revista de Paleontología . 87 (2): 254–68. doi :10.1666/12-080R.1. OCLC 4960242901. S2CID 83864139.
Harlan, R. (1834). "Aviso de huesos fósiles encontrados en la formación Terciaria del Estado de Luisiana". Transacciones de la Sociedad Filosófica Estadounidense . 4 : 397–403. doi :10.2307/1004838. JSTOR 1004838. OCLC 63356837.
Héctor, J. (1881). "Notas sobre los cetáceos de Nueva Zelanda, recientes y fósiles". Transacciones y procedimientos del Instituto de Nueva Zelanda . 13 : 434–37. OCLC 31837919 . Consultado el 1 de julio de 2013 .
Hulbert, Richard C. Jr.; Petkewich, Richard M.; Obispo, Gale A.; Bukry, David; Aleshire, David P. (septiembre de 1998). "Una nueva ballena protocétida del Eoceno medio (Mammalia: Cetacea: Archaeoceti) y biota asociada de Georgia". Revista de Paleontología . 72 (5): 907–927. doi :10.1017/S0022336000027232. JSTOR 1306667. OCLC 4908698029. S2CID 131420545.
Kumar, K.; Sahni, A. (1986). " Remingtonocetus harudiensis , nueva combinación, un arqueoceto del Eoceno medio (Mammalia, Cetacea) del oeste de Kutch, India". Revista de Paleontología de Vertebrados . 6 (4): 326–349. doi :10.1080/02724634.1986.10011629. OCLC 4649653943.
McLeod, SA; Barnes, LG (2008). "Un nuevo género y especie de ballena arqueoceta protocétida del Eoceno (Mammalia, Cetacea) de la llanura costera atlántica" (PDF) . En Wang, Xiaoming; Barnes, Lawrence G. (eds.). Geología y paleontología de vertebrados del oeste y sur de América del Norte . vol. 41, págs. 73–98 . Consultado el 1 de febrero de 2013 . {{cite book}}: |journal=ignorado ( ayuda )
Rosa, Kenneth David (2006). El comienzo de la era de los mamíferos. Prensa JHU. ISBN 978-0-8018-8472-6.
Sahni, Ashok; Mishra, Vijay Prakash (1975). "Vertebrados del Terciario Inferior del oeste de la India". Monografía de la Sociedad Paleontológica de la India . 3 : 1–48. COMO EN B0007AL8UE. OCLC 3566369.
Steeman, YO; Hebsgaard, MB; Fordyce, R. Ewan; Ho, SYW; Rabosky, DL; Nielsen, R.; Rahbek, C.; Glenner, H.; Sorensen, MV; Willerslev, E. (2009). "Radiación de cetáceos existentes impulsada por la reestructuración de los océanos". Biología Sistemática . 58 (6): 573–585. doi :10.1093/sysbio/syp060. PMC 2777972 . PMID 20525610.
Stromer, Ernst (1908). Die Archaeoceti des ägyptischen Eozäns: Beiträge zur paläontologie und geoologie Österreich-Ungarns und des Orients (PDF) . W. Braumüller. OCLC 21174007 . Consultado el 1 de julio de 2013 .
Thewissen, JGM (2002). "Arqueocetos, Arcaicos". En Perrin, William R; Wiirsig, Bernd; Thewissen , JGM (eds.). Enciclopedia de mamíferos marinos . Prensa académica. págs. 36–9. ISBN 978-0-12-551340-1.
Thewissen, JGM ; Hussain, ST (2000). " Attockicetus praecursor , un nuevo cetáceo remingtonocetido de sedimentos marinos del Eoceno de Pakistán". Revista de evolución de los mamíferos . 7 (3): 133–46. doi :10.1023/A:1009458618729. OCLC 362777268. S2CID 5700031.
Thewissen, JGM ; Hussain, ST (1998). "Revisión sistemática de Pakicetidae, Cetacea (Mammalia) del Eoceno temprano y medio de Pakistán e India". Boletín del Museo Carnegie . 34 : 220–38.
Thewissen, JGM ; Madar, SI; Hussain, ST (1996).Ambulocetus natans , un cetáceo del Eoceno (Mammalia) de Pakistán . Correo Forschungsinstitut Senckenberg. vol. 191, págs. 1–86. ISBN 978-3-929907-32-2. OCLC 36463214.
Thewissen, JGM ; Williams, EM; Roe, LJ; Hussain, ST (2001). "Esqueletos de cetáceos terrestres y la relación de las ballenas con los artiodáctilos" (PDF) . Naturaleza . 413 (6853): 277–81. Código Bib :2001Natur.413..277T. doi :10.1038/35095005. OCLC 118116179. PMID 11565023. S2CID 4416684 . Consultado el 1 de febrero de 2013 .
Trivedy, AN; Satsangi, PP (1984). "Un nuevo arqueoceto (ballena) del Eoceno de la India". Resúmenes del 27º Congreso Geológico Internacional, Moscú . 1 : 322–23.
Es cierto, FW (1908). "El cetáceo fósil Dorudon serratus Gibbes". Boletín del Museo de Zoología Comparada . 52 (4): 5–78. OCLC 355813868. OL 19219818M . Consultado el 1 de julio de 2013 .
Uhen, Mark D (1998). "Nuevo protocétido (Mammalia, Cetacea) de la Formación Cook Mountain del Eoceno medio tardío de Luisiana". Revista de Paleontología de Vertebrados . 18 (3): 664–8. doi :10.1080/02724634.1998.10011093. JSTOR 4523937. OCLC 204612030.
Uhen, Mark D. (2005). "Un nuevo género y especie de ballena arqueoceto de Mississippi". Geología del Sureste . 43 (3): 157–72.
Uhen, MD (2008). "Nuevas ballenas protocétidas de Alabama y Mississippi, y un nuevo clado de cetáceos, Pelagiceti". Revista de Paleontología de Vertebrados . 28 (3): 589–593. doi :10.1671/0272-4634(2008)28[589:NPWFAA]2.0.CO;2. S2CID 86326007.
"MD Uhen 2008". Obras fósiles .
Uhen, Mark D; Gingerich, Philip D (enero de 2001). "Nuevo género de arqueoceto dorudontino (Cetacea) del Eoceno medio a tardío de Carolina del Sur" (PDF) . Ciencia de los mamíferos marinos . 17 (1): 1–34. doi :10.1111/j.1748-7692.2001.tb00979.x. hdl : 2027.42/73005 . OCLC 204061291.
Uhen, Mark D.; Pyenson, Nicolás D .; Devries, Thomas J.; Urbina, Mario; Renne, Paul R. (2011). "Nuevas Ballenas del Eoceno Medio de la Cuenca de Pisco del Perú". Revista de Paleontología . 85 (5): 955–69. doi :10.1666/10-162.1. hdl :10088/17509. OCLC 802202947. S2CID 115130412.
Oeste, Robert M (1980). "Ensamblaje de grandes mamíferos del Eoceno medio con afinidades de Tethyan, región de Ganda Kas, Pakistán". Revista de Paleontología . 54 (3): 508–533. JSTOR 1304193. OCLC 4899161959.
enlaces externos
Medios relacionados con Archaeoceti en Wikimedia Commons




{kind=link}