Plasmodium es un género de eucariotas unicelularesque son parásitos obligados de vertebrados e insectos . Los ciclos de vida delas especies de Plasmodium implican el desarrollo en un insecto huésped hematófago que luego inyecta parásitos en un huésped vertebrado durante una comida de sangre. Los parásitos crecen dentro de un tejido corporal vertebrado (a menudo el hígado) antes de ingresar al torrente sanguíneo para infectar los glóbulos rojos . La consiguiente destrucción de los glóbulos rojos del huésped puede provocar malaria . Durante esta infección, algunos parásitos son recogidos por un insecto hematófago ( mosquitos en la mayoría de los casos), continuando el ciclo de vida. [1]
Plasmodium es un miembro del filo Apicomplexa , un gran grupo de eucariotas parásitos. Dentro de Apicomplexa, Plasmodium pertenece al orden Haemosporida y a la familia Plasmodiidae . Se han descrito más de 200 especies de Plasmodium , muchas de las cuales se han subdividido en 14 subgéneros según la morfología del parásito y la gama de hospedadores. Las relaciones evolutivas entre las diferentes especies de Plasmodium no siempre siguen límites taxonómicos; algunas especies que son morfológicamente similares o que infectan al mismo hospedador resultan estar lejanamente relacionadas.
Las especies de Plasmodium se distribuyen globalmente dondequiera que se encuentren huéspedes adecuados. Los huéspedes insectos son con mayor frecuencia mosquitos de los géneros Culex y Anopheles . Los huéspedes vertebrados incluyen reptiles, aves y mamíferos. Los parásitos de Plasmodium fueron identificados por primera vez a fines del siglo XIX por Charles Laveran . A lo largo del siglo XX, se descubrieron y clasificaron muchas otras especies en varios huéspedes, incluidas cinco especies que infectan regularmente a los humanos: P. vivax , P. falciparum , P. malariae , P. ovale y P. knowlesi . P. falciparum es, con mucho, el más letal en humanos, lo que resulta en cientos de miles de muertes por año. Se han desarrollado varios medicamentos para tratar la infección por Plasmodium ; sin embargo, los parásitos han desarrollado resistencia a cada medicamento desarrollado.
Aunque el parásito también puede infectar a las personas a través de transfusiones de sangre , esto es muy poco frecuente y el Plasmodium no se puede transmitir de persona a persona. Algunas subespecies de Plasmodium son parásitos intracelulares obligados .

El género Plasmodium está formado por todos los eucariotas del filo Apicomplexa que experimentan el proceso de replicación asexual de merogonia dentro de los glóbulos rojos del huésped y producen el pigmento cristalino hemozoína como subproducto de la digestión de la hemoglobina del huésped . [2] Las especies de Plasmodium contienen muchas características que son comunes a otros eucariotas, y algunas que son exclusivas de su filo o género. El genoma de Plasmodium está separado en 14 cromosomas contenidos en el núcleo . Los parásitos de Plasmodium mantienen una única copia de su genoma durante gran parte de su ciclo de vida, duplicando el genoma solo para un breve intercambio sexual dentro del intestino medio del insecto huésped. [3] Unido al núcleo está el retículo endoplasmático (RE), que funciona de manera similar al RE en otros eucariotas. Las proteínas se transportan desde el RE al aparato de Golgi , que generalmente consta de un único compartimento limitado por la membrana en Apicomplexans. [4] Desde aquí, las proteínas son transportadas a diversos compartimentos celulares o a la superficie celular. [4]
Al igual que otros apicomplejos, las especies de Plasmodium tienen varias estructuras celulares en el extremo apical del parásito que sirven como orgánulos especializados para secretar efectores en el huésped. Los más destacados son las roptrias bulbosas que contienen proteínas del parásito implicadas en la invasión de la célula huésped y la modificación del huésped una vez dentro. [5] Adyacentes a las roptrias hay estructuras más pequeñas denominadas micronemas que contienen proteínas del parásito necesarias para la motilidad, así como para reconocer y unirse a las células huésped. [6] Distribuidas por todo el parásito hay vesículas secretoras llamadas gránulos densos que contienen proteínas del parásito implicadas en la modificación de la membrana que separa al parásito del huésped, denominada vacuola parasitófora . [6]
Las especies de Plasmodium también contienen dos grandes orgánulos unidos a membranas de origen endosimbiótico , la mitocondria y el apicoplasto , ambos desempeñan papeles clave en el metabolismo del parásito . A diferencia de las células de mamíferos que contienen muchas mitocondrias, las células de Plasmodium contienen una única mitocondria grande que coordina su división con la de la célula de Plasmodium . [7] Al igual que en otros eucariotas, la mitocondria de Plasmodium es capaz de generar energía en forma de ATP a través del ciclo del ácido cítrico ; sin embargo, esta función solo es necesaria para la supervivencia del parásito en el huésped insecto, y no es necesaria para el crecimiento de los glóbulos rojos. [7] Un segundo orgánulo, el apicoplasto, se deriva de un evento de endosimbiosis secundario , en este caso la adquisición de un alga roja por parte del ancestro de Plasmodium . [8] El apicoplasto participa en la síntesis de varios precursores metabólicos, incluidos ácidos grasos , isoprenoides , grupos de hierro-azufre y componentes de la vía de biosíntesis del hemo . [9]


El ciclo de vida de Plasmodium involucra varias etapas distintas en los huéspedes insectos y vertebrados . Los parásitos generalmente se introducen en un huésped vertebrado por la picadura de un insecto huésped (generalmente un mosquito, con la excepción de algunas especies de Plasmodium de reptiles). [10] Los parásitos primero infectan el hígado u otro tejido, donde experimentan una sola gran ronda de replicación antes de salir de la célula huésped para infectar los eritrocitos . [11] En este punto, algunas especies de Plasmodium de primates pueden formar una etapa latente de larga duración llamada hipnozoito, [12] que puede permanecer en el hígado durante más de un año. [13] Sin embargo, para la mayoría de las especies de Plasmodium , los parásitos en las células hepáticas infectadas son solo lo que se llama merozoitos. Después de emerger del hígado, ingresan a los glóbulos rojos, como se explicó anteriormente. Luego pasan por ciclos continuos de infección de eritrocitos, mientras que un pequeño porcentaje de parásitos se diferencian en una etapa sexual llamada gametocito, que es recogido por un insecto huésped que se alimenta de sangre. En algunos huéspedes, la invasión de eritrocitos por especies de Plasmodium puede provocar una enfermedad llamada malaria. A veces puede ser grave y rápidamente seguida de la muerte del huésped (por ejemplo, P. falciparum en humanos). En otros huéspedes, la infección por Plasmodium puede ser aparentemente asintomática. [10]
Incluso cuando los seres humanos padecen infecciones plasmódicas subclínicas, puede haber, no obstante, una gran cantidad de parásitos multiplicándose ocultos, en particular, en el bazo y la médula ósea. Esto es así en el caso de P. vivax . Se cree que estos parásitos ocultos (además de los hipnozoítos) son el origen de los casos recurrentes de malaria por P. vivax . [14]
.jpg/1280px-Malaria_Sporozoites_(16657642929).jpg)
Dentro de los glóbulos rojos, los merozoitos crecen primero hasta una forma en forma de anillo y luego hasta una forma más grande llamada trofozoito . Luego, los trofozoitos maduran hasta convertirse en esquizontes que se dividen varias veces para producir nuevos merozoitos. El glóbulo rojo infectado finalmente estalla, lo que permite que los nuevos merozoitos viajen dentro del torrente sanguíneo para infectar nuevos glóbulos rojos. La mayoría de los merozoitos continúan este ciclo replicativo, sin embargo, algunos merozoitos, al infectar glóbulos rojos, se diferencian en formas sexuales masculinas o femeninas llamadas gametocitos. Estos gametocitos circulan en la sangre hasta que son absorbidos cuando un mosquito se alimenta del huésped vertebrado infectado, absorbiendo sangre que incluye los gametocitos. [11]
En el mosquito, los gametocitos se desplazan junto con la sangre ingerida hasta el intestino medio del mosquito. Allí, los gametocitos se convierten en gametos masculinos y femeninos que se fecundan entre sí, formando un cigoto . Luego, los cigotos se convierten en una forma móvil llamada oocineto , que penetra la pared del intestino medio. Al atravesar la pared del intestino medio, el oocineto se incrusta en la membrana exterior del intestino y se convierte en un ooquiste. Los ooquistes se dividen muchas veces para producir una gran cantidad de pequeños esporozoitos alargados . Estos esporozoitos migran a las glándulas salivales del mosquito, donde pueden inyectarse en la sangre del siguiente huésped al que pica el mosquito, repitiendo el ciclo. [11]

Plasmodium pertenece al filo Apicomplexa , un grupo taxonómico de parásitos unicelulares con orgánulos secretores característicos en un extremo de la célula. [15] Dentro de Apicomplexa, Plasmodium está dentro del orden Haemosporida , un grupo que incluye todos los apicomplejos que viven dentro de las células sanguíneas. [16] Basado en la presencia del pigmento hemozoína y el método de reproducción asexual , el orden se divide en cuatro familias, de las cuales Plasmodium está en la familia Plasmodiidae . [17]
El género Plasmodium consta de más de 200 especies, generalmente descritas en función de su apariencia en frotis de sangre de vertebrados infectados. [18] Estas especies se han categorizado en función de su morfología y rango de hospedadores en 14 subgéneros: [17]
Las especies que infectan a monos y simios, con excepción de P. falciparum y P. reichenowi (que juntas forman el subgénero Laverania ), se clasifican en el subgénero Plasmodium . Los parásitos que infectan a otros mamíferos, incluidos algunos primates ( lémures y otros), se clasifican en el subgénero Vinckeia . Los cinco subgéneros Bennettinia , Giovannolaia , Haemamoeba , Huffia y Novyella contienen las especies palúdicas aviares conocidas. [19] Los subgéneros restantes: Asiamoeba , Carinamoeba , Lacertamoeba , Ophidiella , Paraplasmodium y Sauramoeba contienen los diversos grupos de parásitos que infectan a los reptiles. [20]
Estudios más recientes de especies de Plasmodium utilizando métodos moleculares han implicado que la evolución del grupo no ha seguido perfectamente la taxonomía. [2] Muchas especies de Plasmodium que son morfológicamente similares o infectan a los mismos huéspedes resultan estar solo distantemente relacionadas. [21] En la década de 1990, varios estudios buscaron evaluar las relaciones evolutivas de las especies de Plasmodium comparando el ARN ribosómico y un gen de proteína de superficie de varias especies, encontrando que el parásito humano P. falciparum estaba más estrechamente relacionado con los parásitos aviares que con otros parásitos de primates. [17] Sin embargo, estudios posteriores que muestrearon más especies de Plasmodium encontraron que los parásitos de los mamíferos forman un clado junto con el género Hepatocystis , mientras que los parásitos de las aves o lagartijas parecen formar un clado separado con relaciones evolutivas que no siguen los subgéneros: [17] [22]
Las estimaciones sobre la divergencia de los diferentes linajes de Plasmodium han variado considerablemente. Las estimaciones sobre la divergencia del orden Haemosporida varían entre hace unos 16,2 millones y 100 millones de años. [17] Ha habido un interés particular en datar la divergencia del parásito humano P. falciparum de otros linajes de Plasmodium debido a su importancia médica. Para ello, las fechas estimadas varían entre hace 110.000 y 2,5 millones de años. [17]
Las especies de Plasmodium se distribuyen por todo el mundo. Todas son parásitas y deben pasar de un hospedador vertebrado a un hospedador insecto para completar su ciclo de vida. Las diferentes especies de Plasmodium presentan diferentes rangos de hospedadores: algunas especies se limitan a un solo hospedador vertebrado e insecto, mientras que otras pueden infectar a varias especies de vertebrados y/o insectos.
_species_by_country_of_origin_for_imported_cases_to_non-endemic_countries.png/1280px-Relative_incidence_of_Plasmodium_(malaria)_species_by_country_of_origin_for_imported_cases_to_non-endemic_countries.png)
Los parásitos Plasmodium se han descrito en una amplia gama de huéspedes vertebrados, incluidos reptiles, aves y mamíferos. [24] Si bien muchas especies pueden infectar a más de un huésped vertebrado, generalmente son específicas de una de estas clases (como las aves). [24]
Los humanos son infectados principalmente por cinco especies de Plasmodium , y la abrumadora mayoría de las enfermedades graves y muertes son causadas por Plasmodium falciparum . [25] Algunas especies que infectan a los humanos también pueden infectar a otros primates, y las zoonosis de ciertas especies (por ejemplo, P. knowlesi ) de otros primates a los humanos son comunes. [25] Los primates no humanos también contienen una variedad de especies de Plasmodium que generalmente no infectan a los humanos. Algunas de estas pueden causar enfermedades graves en primates, mientras que otras pueden permanecer en el huésped durante períodos prolongados sin causar enfermedades. [26] Muchos otros mamíferos también son portadores de especies de Plasmodium , como una variedad de roedores , ungulados y murciélagos . Nuevamente, algunas especies de Plasmodium pueden causar enfermedades graves en algunos de estos huéspedes, mientras que muchas parecen no hacerlo. [27]
Más de 150 especies de Plasmodium infectan a una amplia variedad de aves. En general, cada especie de Plasmodium infecta a una o varias especies de aves. [28] Los parásitos de Plasmodium que infectan a las aves tienden a persistir en un huésped determinado durante años o durante toda la vida del huésped, aunque en algunos casos las infecciones por Plasmodium pueden provocar una enfermedad grave y una muerte rápida. [29] [30] A diferencia de las especies de Plasmodium que infectan a los mamíferos, las que infectan a las aves están distribuidas por todo el mundo. [28]
Las especies de varios subgéneros de Plasmodium infectan a diversos reptiles . Los parásitos de Plasmodium se han descrito en la mayoría de las familias de lagartijas y, al igual que los parásitos aviares, están distribuidos por todo el mundo. [31] Nuevamente, los parásitos pueden causar una enfermedad grave o ser aparentemente asintomáticos dependiendo del parásito y del huésped. [31]
A lo largo de los años se han desarrollado varios fármacos para controlar la infección por Plasmodium en huéspedes vertebrados, particularmente en humanos. La quinina se utilizó como un antipalúdico de primera línea desde el siglo XVII hasta que surgió una resistencia generalizada a principios del siglo XX. [32] La resistencia a la quinina estimuló el desarrollo de una amplia gama de medicamentos antipalúdicos a lo largo del siglo XX, entre ellos la cloroquina , el proguanil , la atovacuona , la sulfadoxina/pirimetamina , la mefloquina y la artemisinina . [32] En todos los casos, han surgido parásitos resistentes a un fármaco determinado en unas pocas décadas desde el despliegue de los fármacos. [32] Para combatir esto, los fármacos antipalúdicos se utilizan con frecuencia en combinación, y las terapias combinadas con artemisinina son actualmente el estándar de oro para el tratamiento. [33] En general, los fármacos antipalúdicos se dirigen a las etapas de vida de los parásitos Plasmodium que residen dentro de los glóbulos rojos de los vertebrados, ya que estas son las etapas que tienden a causar enfermedades. [34] Sin embargo, se están desarrollando medicamentos dirigidos a otras etapas del ciclo de vida del parásito con el fin de prevenir la infección en viajeros y prevenir la transmisión de las etapas sexuales a los insectos huéspedes. [35]
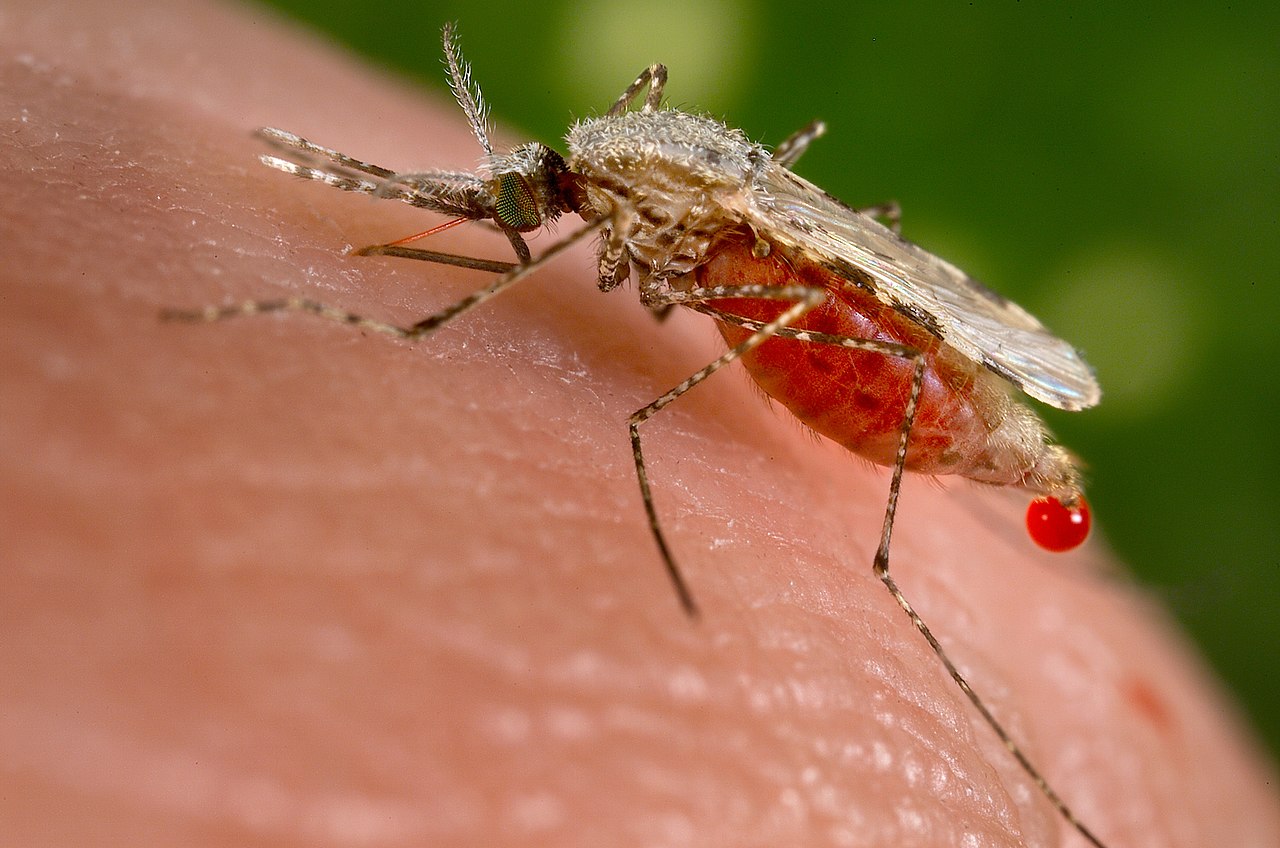
Además de un huésped vertebrado, todas las especies de Plasmodium también infectan a un insecto chupador de sangre , generalmente un mosquito (aunque algunos parásitos que infectan a los reptiles son transmitidos por flebótomos ). Los mosquitos de los géneros Culex , Anopheles , Culiseta , Mansonia y Aedes actúan como insectos huéspedes para varias especies de Plasmodium . Los mejor estudiados de estos son los mosquitos Anopheles que albergan los parásitos Plasmodium de la malaria humana, así como los mosquitos Culex que albergan las especies de Plasmodium que causan malaria en aves. Solo los mosquitos hembra están infectados con Plasmodium , ya que solo ellos se alimentan de la sangre de huéspedes vertebrados. [36] Diferentes especies afectan a sus huéspedes insectos de manera diferente. A veces, los insectos infectados con Plasmodium tienen una esperanza de vida reducida y una capacidad reducida para producir descendencia. [37] Además, algunas especies de Plasmodium parecen hacer que los insectos prefieran picar a huéspedes vertebrados infectados en lugar de a huéspedes no infectados. [37] [38] [39]
Charles Louis Alphonse Laveran describió por primera vez los parásitos en la sangre de pacientes con malaria en 1880. [40] Llamó al parásito Oscillaria malariae . [40] En 1885, los zoólogos Ettore Marchiafava y Angelo Celli reexaminaron el parásito y lo denominaron miembro de un nuevo género, Plasmodium , llamado así por la semejanza con las células multinucleadas de los mohos mucilaginosos del mismo nombre. [41] [notas 1] El hecho de que varias especies puedan estar implicadas en causar diferentes formas de malaria fue reconocido por primera vez por Camillo Golgi en 1886. [40] Poco después, Giovanni Batista Grassi y Raimondo Filetti nombraron a los parásitos que causaban dos tipos diferentes de malaria humana Plasmodium vivax y Plasmodium malariae . [40] En 1897, William Welch identificó y nombró a Plasmodium falciparum . A esto le siguió el reconocimiento de las otras dos especies de Plasmodium que infectan a los humanos: Plasmodium ovale (1922) y Plasmodium knowlesi (identificado en macacos de cola larga en 1931; en humanos en 1965). [40] La contribución de los insectos huéspedes al ciclo de vida de Plasmodium fue descrita en 1897 por Ronald Ross y en 1899 por Giovanni Batista Grassi, Amico Bignami y Giuseppe Bastianelli . [40]
En 1966, Cyril Garnham propuso separar Plasmodium en nueve subgéneros basándose en la especificidad del hospedador y la morfología del parásito. [18] Esto incluía cuatro subgéneros que A. Corradetti había propuesto previamente para las especies de Plasmodium que infectan a las aves en 1963. [42] [19] Sam R. Telford amplió este esquema en 1988 cuando reclasificó los parásitos de Plasmodium que infectan a los reptiles, añadiendo cinco subgéneros. [20] [18] En 1997, G. Valkiunas reclasificó las especies de Plasmodium que infectan a las aves añadiendo un quinto subgénero: Bennettinia . [19] [43]

