Aunque un estudio genético indica que R1a se originó hace 25.000 [2] años, su subclado M417 (R1a1a1) se diversificó hace aproximadamente 5.800 años. [4] El lugar de origen del subclado juega un papel en el debate sobre los orígenes de los protoindoeuropeos .
La mutación SNP R-M420 fue descubierta después de R-M17 (R1a1a), lo que resultó en una reorganización del linaje, estableciendo en particular un nuevo paragrupo (designado R-M420*) para los linajes relativamente raros que no están en la rama R-SRY10831.2 (R1a1) que conduce a R-M17.
Orígenes
Orígenes de R1a
Se estima que la divergencia genética de R1a (M420) ocurrió hace 25.000 [2] años, que es el momento del último máximo glacial . Un estudio de 2014 realizado por Peter A. Underhill et al., utilizando 16.244 individuos de más de 126 poblaciones de toda Eurasia, concluyó que había "un caso convincente para el Medio Oriente, posiblemente cerca del actual Irán, como el origen geográfico de hg R1a". [2] El registro de ADN antiguo ha mostrado el primer R1a durante el Mesolítico en cazadores-recolectores orientales (de Europa del Este, hace unos 13.000 años), [5] [6] y el caso más temprano de R* entre los antiguos euroasiáticos del norte del Paleolítico superior , [7] de donde los cazadores-recolectores orientales derivan predominantemente su ascendencia. [8]
Diversificación de R1a1a1 (M417) y migraciones antiguas
Orígenes de R1a (Underhill 2009; [3] Orígenes de R1a1a (Pamjav et al. 2012); posible migración de R1a a la costa báltica; y la expansión más antigua y la frecuencia más alta de R1a1a (Underhill et al. 2014)
Según Underhill et al. (2014), el subclado R1a-M417 aguas abajo se diversificó en Z282 y Z93 hace unos 5.800 años "en las proximidades de Irán y el este de Turquía". [4] [nota 1] Aunque R1a aparece como un haplogrupo del cromosoma Y entre hablantes de varias lenguas como el eslavo y el indoiraní , la cuestión de los orígenes de R1a1a es relevante para el debate en curso sobre el urheimat del pueblo protoindoeuropeo , y también puede ser relevante para los orígenes de la civilización del valle del Indo . R1a muestra una fuerte correlación con las lenguas indoeuropeas del sur y oeste de Asia , Europa central y oriental y Escandinavia [10] [3] siendo más frecuente en Europa del Este , Asia Central y Asia del Sur . En Europa, Z282 es prevalente particularmente mientras que en Asia domina Z93. La conexión entre el ADN-Y R-M17 y la difusión de las lenguas indoeuropeas fue observada por primera vez por T. Zerjal y sus colegas en 1999. [11]
Relación indoeuropea
Propuesta de dispersión esteparia de R1a1a
Semino et al. (2000) propusieron orígenes ucranianos y una propagación postglacial del haplogrupo R1a1 durante el Máximo Glacial Tardío , posteriormente magnificada por la expansión de la cultura Kurgan hacia Europa y hacia el este. [12] Spencer Wells propone orígenes de Asia Central, sugiriendo que la distribución y la edad de R1a1 apuntan a una antigua migración correspondiente a la propagación del pueblo Kurgan en su expansión desde la estepa euroasiática . [13] Según Pamjav et al. (2012), R1a1a se diversificó en las estepas euroasiáticas o en la región de Oriente Medio y el Cáucaso:
Asia interior y central es una zona de superposición para los linajes R1a1-Z280 y R1a1-Z93 [lo que] implica que una zona de diferenciación temprana de R1a1-M198 posiblemente ocurrió en algún lugar dentro de las estepas euroasiáticas o la región del Medio Oriente y el Cáucaso, ya que se encuentran entre el sur de Asia y Europa central y oriental. [14]
Tres estudios genéticos realizados en 2015 dieron apoyo a la teoría kurganiana de Gimbutas en relación con el Urheimat indoeuropeo . Según esos estudios, los haplogrupos R1b y R1a, ahora los más comunes en Europa (R1a también es común en el sur de Asia) se habrían expandido desde las estepas póntico-caspias, junto con las lenguas indoeuropeas; también detectaron un componente autosómico presente en los europeos modernos que no estaba presente en los europeos neolíticos, que se habría introducido con los linajes paternos R1b y R1a, así como con las lenguas indoeuropeas. [15] [16] [17]
Silva et al. (2017) observaron que R1a en el sur de Asia "probablemente se propagó a partir de un único grupo de origen de Asia central , parece haber al menos tres y probablemente más clados fundadores de R1a dentro del subcontinente indio , en consonancia con múltiples olas de llegada". [18] Según Martin P. Richards, coautor de Silva et al. (2017), la prevalencia de R1a en la India fue "una prueba muy poderosa de una migración sustancial de la Edad de Bronce desde Asia central que probablemente trajo hablantes indoeuropeos a la India". [19] [nota 2]
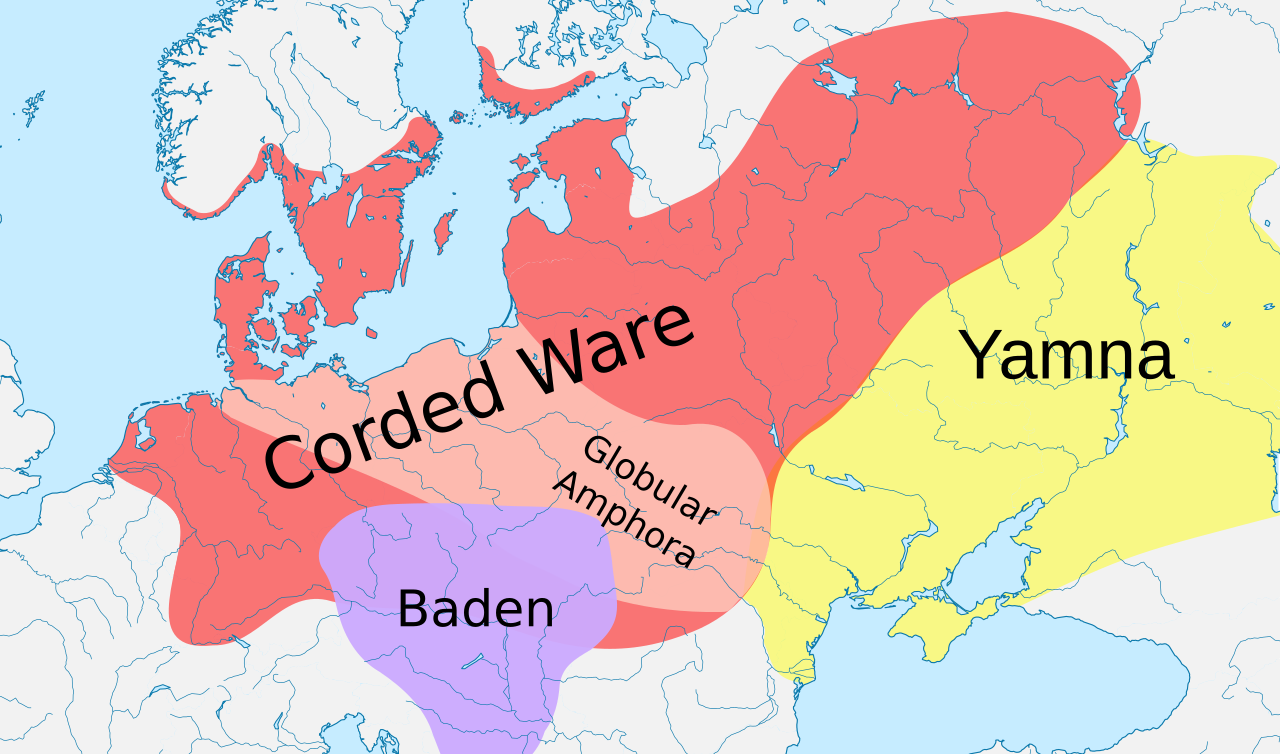
Posibles orígenes de Yamnaya o cerámica cordada
Período Neolítico Medio Europeo. Cultura de la Cerámica Peine, c. 4200 – c. 2000 a. C.Cultura de la cerámica cordada (c. 2900 – c. 2350 a. C.)
David Anthony considera que la cultura Yamnaya es la Urheimat indoeuropea . [20] [21] Según Haak et al. (2015), una migración masiva de la cultura Yamnaya hacia el norte tuvo lugar alrededor del 2500 a. C., lo que representa el 75% de la ascendencia genética de la cultura Corded Ware , y señala que R1a y R1b pueden haberse "extendido a Europa desde el este después del 3000 a. C.". [22] Sin embargo, sus siete muestras Yamnaya pertenecían al subclado R1b-M269 , [22] pero no se ha encontrado R1a1a en sus muestras Yamnaya. Esto plantea la pregunta de dónde vino el R1a1a en la cultura Corded Ware, si no era de la cultura Yamnaya. [23]
Según Marc Haber, la ausencia del haplogrupo R1a-M458 en Afganistán no respalda un origen de estepa póntico-caspia para los linajes R1a en las poblaciones modernas de Asia Central. [24]
El arqueólogo Barry Cunliffe ha dicho que la ausencia del haplogrupo R1a en los especímenes Yamnaya es una debilidad importante en la propuesta de Haak de que R1a tiene un origen Yamnaya. [26]
Semenov y Bulat (2016) argumentan a favor de un origen Yamnaya de R1a1a en la cultura de cerámica cordada, y señalan que varias publicaciones apuntan a la presencia de R1a1 en la cultura de cerámica peine . [27] [nota 3]
Orígenes propuestos del sur de Asia
Kivisild et al. (2003) han propuesto Asia meridional o occidental , [28] [nota 4] mientras que Mirabal et al. (2009) ven apoyo tanto para Asia meridional como central. [10] Sengupta et al. (2006) han propuesto orígenes indios. [29] Thanseem et al. (2006) han propuesto Asia meridional o central. [30] Sahoo et al. (2006) han propuesto Asia meridional o occidental. [31] Thangaraj et al. (2010) también han propuesto un origen en el sur de Asia. [32] Sharma et al. (2009) teorizan la existencia de R1a en la India más allá de los 18.000 años hasta posiblemente 44.000 años de origen. [1]
Varios estudios realizados entre 2006 y 2010 concluyeron que las poblaciones del sur de Asia tienen la mayor diversidad de STR dentro de R1a1a, [33] [34] [10] [3] [1] [35] y dataciones TMRCA posteriores más antiguas . [nota 5] R1a1a está presente tanto entre las castas superiores ( brahmanes ) como entre las inferiores, y aunque la frecuencia es mayor entre las castas brahmanes, las dataciones TMRCA más antiguas del haplogrupo R1a se dan en la tribu Saharia , una casta programada de la región Bundelkhand de la India central . [1] [35]
A partir de estos hallazgos, algunos investigadores argumentaron que el R1a1a se originó en el sur de Asia, [34] [1] [nota 6] excluyendo un influjo genético más reciente, aunque menor, de migrantes indoeuropeos en regiones del noroeste como Afganistán, Baluchistán, Punjab y Cachemira. [34] [33] [3] [nota 7]
La conclusión de que R1a se originó en la India ha sido cuestionada por investigaciones más recientes, [18] [37] [nota 8] que ofrecen pruebas de que R1a llegó a la India con múltiples olas de migración. [18] [38]
Orígenes propuestos de Transcaucasia y Asia occidental y posible influencia en la civilización del valle del Indo
Haak et al. (2015) descubrieron que parte de la ascendencia Yamnaya se derivó de Oriente Medio y que las técnicas neolíticas probablemente llegaron a la cultura Yamnaya desde los Balcanes . [nota 9] La cultura Rössen (4.600–4.300 a. C.), que estaba situada en Alemania y es anterior a la cultura Corded Ware, un antiguo subclado de R1a, concretamente L664, aún se puede encontrar. [nota 10]
Parte de la ascendencia genética del sur de Asia se deriva de poblaciones del oeste de Eurasia, y algunos investigadores han insinuado que Z93 puede haber llegado a la India a través de Irán [40] y haberse expandido allí durante la civilización del valle del Indo . [2] [41]
Mascarenhas et al. (2015) propusieron que las raíces de Z93 se encuentran en Asia occidental, y propusieron que "Z93 y L342.2 se expandieron en dirección sureste desde Transcaucasia hasta el sur de Asia ", [40] señalando que dicha expansión es compatible con "los registros arqueológicos de expansión hacia el este de las poblaciones de Asia occidental en el cuarto milenio a. C. que culminaron en las llamadas migraciones Kura-Araxes en el período posterior a Uruk IV ". [40] Sin embargo, Lazaridis señaló que la muestra I1635 de Lazaridis et al. (2016), su muestra armenia Kura-Araxes, contenía el haplogrupo Y R1 b 1-M415(xM269) [nota 11] (también llamado R1b1a1b-CTS3187). [42] [ ¿ fuente poco confiable? ]
Según Underhill et al. (2014), la diversificación de Z93 y la "urbanización temprana dentro del valle del Indo... ocurrieron [hace 5.600 años] y la distribución geográfica de R1a-M780 (Figura 3d [nota 12] ) puede reflejar esto". [2] [nota 13] Poznik et al. (2016) señalan que se produjeron "expandciones sorprendentes" dentro de R1a-Z93 hace unos 4.500-4.000 años, lo que "preda por unos pocos siglos el colapso de la civilización del valle del Indo". [41] [nota 14]
Sin embargo, según Narasimhan et al. (2018), los pastores esteparios son una fuente probable de R1a en la India. [44] [nota 15]
Filogenia
El árbol genealógico R1a ahora tiene tres niveles principales de ramificación, con el mayor número de subclados definidos dentro de la rama dominante y mejor conocida, R1a1a (que se encontrará con varios nombres como "R1a1" en la literatura relativamente reciente pero no en la más reciente).
Topología
La topología de R1a es la siguiente (códigos [entre corchetes] códigos no isogg): [9] [45] [ verificación necesaria ] [46] [2] [47] Tatiana et al. (2014) "el rápido proceso de diversificación de K-M526 probablemente ocurrió en el sudeste asiático , con posteriores expansiones hacia el oeste de los ancestros de los haplogrupos R y Q ". [48]
R1a1a1b1a1 [46] [El código topológico antiguo es R1a1a1b*, que está desactualizado y puede generar cierta confusión.] [49] (M458) [46] [49] [R1a1a1g] (M458) [47]
El gen R1a se distingue por varios marcadores únicos, incluida la mutación M420. Es un subclado del haplogrupo R-M173 (anteriormente llamado R1). El gen R1a tiene los subclados hermanos Haplogrupo R1b -M343 y el paragrupo R-M173*.
R-M420 (R1a)
R-M420, definido por la mutación M420, tiene dos ramas: R-SRY1532.2, definido por la mutación SRY1532.2, que constituye la gran mayoría; y R-M420*, el paragrupo , definido como positivo para M420 pero negativo para SRY1532.2. (En el esquema de 2002, esta minoría negativa para SRY1532.2 era una parte del grupo relativamente raro clasificado como el paragrupo R1*). Las mutaciones que se entiende que son equivalentes a M420 incluyen M449, M511, M513, L62 y L63. [3] [51]
Este paragrupo se conoce actualmente como R1a2 (R-YP4141) y tiene dos ramificaciones: R1a2a (R-YP5018) y R1a2b (R-YP4132).
R-SRY1532.2 (R1a1)
R1a1 se define como SRY1532.2 o SRY10831.2 (que siempre incluye SRY10831.2, M448, L122, M459 y M516 [3] [52] ). Esta familia de linajes está dominada por M17 y M198. Por el contrario, el paragrupo R-SRY1532.2* carece de los marcadores M17 o M198.
El paragrupo R-SRY1532.2* es aparentemente menos raro que el R1*, pero aún así relativamente inusual, aunque se ha probado en más de un estudio. Underhill et al. (2009) informaron 1/51 en Noruega , 3/305 en Suecia , 1/57 macedonios griegos , 1/150 iraníes, 2/734 armenios étnicos y 1/141 kabardianos . [3] Sahoo et al. (2006) informaron R-SRY1532.2* para 1/15 muestras de Rajput de Himachal Pradesh . [34]
R-M17/M198 (R1a1a)
Los siguientes SNP están asociados con R1a1a:
R-M417 (R1a1a1)
R1a1a1 (R-M417) es el subclado más ampliamente encontrado, en dos variaciones que se encuentran respectivamente en Europa (R1a1a1b1 (R-Z282) ([R1a1a1a*] (R-Z282) (Underhill 2014) [2] ) y Asia central y meridional (R1a1a1b2 (R-Z93) ([R1a1a2*] (R-Z93) Underhill 2014) [2] ).
R-Z282 (R1a1a1b1a) (Europa del Este)
Este gran subclado parece abarcar la mayor parte del R1a1a encontrado en Europa. [14]
R1a1a1b1a [R1a1a1a* (Underhill (2014))] (R-Z282*) se presenta en el norte de Ucrania, Bielorrusia y Rusia con una frecuencia de alrededor del 20 %. [2]
R1a1a1b1a3 [R1a1a1a1 (Underhill (2014))] (R-Z284) se encuentra en el noroeste de Europa y alcanza un máximo de alrededor del 20 % en Noruega. [2]
R1a1a1c (M64.2, M87, M204) es aparentemente raro: se encontró en 1 de 117 machos tipificados en el sur de Irán. [53]
R-M458 (R1a1a1b1a1)
Distribución de frecuencias de R-M458
R-M458 es un SNP principalmente eslavo , caracterizado por su propia mutación, y primero fue llamado grupo N. Underhill et al. (2009) encontraron que estaba presente en poblaciones europeas modernas aproximadamente entre la cuenca del Rin y los Montes Urales y lo rastrearon hasta "un efecto fundador que ... cae en el período Holoceno temprano, 7,9 ± 2,6 KYA". (Velocidades de Zhivotovsky, sobrevaloradas 3x) [3] M458 se encontró en un esqueleto de un campo de tumbas del siglo XIV en Usedom , Mecklemburgo-Pomerania Occidental, Alemania. [54] El artículo de Underhill et al. (2009) también informa una frecuencia sorprendentemente alta de M458 en algunas poblaciones del Cáucaso del Norte (18% entre Ak Nogai , [55] 7,8% entre Qara Nogai y 3,4% entre Abazas ). [56]
R-L260 (R1a1a1b1a1a)
R1a1a1b1a1a (R-L260), comúnmente conocido como eslavo occidental o polaco , es un subclado del grupo parental más grande R-M458, y fue identificado por primera vez como un grupo STR por Pawlowski et al. 2002. En 2010 se verificó que era un haplogrupo identificado por su propia mutación (SNP). [57] Aparentemente representa alrededor del 8% de los hombres polacos, lo que lo convierte en el subclado más común en Polonia. Fuera de Polonia es menos común. [58] Además de Polonia, se encuentra principalmente en la República Checa y Eslovaquia , y se considera "claramente eslavo occidental". Se estima que el ancestro fundador de R-L260 vivió entre 2000 y 3000 años atrás, es decir, durante la Edad del Hierro , con una importante expansión poblacional hace menos de 1500 años. [59]
R-M334
Underhill et al. (2009) encontraron R-M334 ([R1a1a1g1], [47] un subclado de [R1a1a1g] (M458) [47] cq R1a1a1b1a1 (M458) [46] ) solo en un hombre estonio y puede definir un clado muy recientemente fundado y pequeño. [3]
R1a1a1b1a2 (S466/Z280, S204/Z91)
R1a1a1b1a2b3* (grupo K de Gwozdz)
R1a1a1b1a2b3* (M417+, Z645+, Z283+, Z282+, Z280+, CTS1211+, CTS3402, Y33+, CTS3318+, Y2613+) (Grupo K de Gwozdz) [45] [ verificación necesaria ] es un grupo basado en STR que es R-M17(xM458). Este grupo es común en Polonia, pero no exclusivo de ese país. [59]
R1a1a1b1a2b3a (R-L365)
R1a1a1b1a2b3a (R-L365) [46] se denominó en un principio Cluster G. [ cita requerida ]
R-Z93* o R1a1a1b2* (R1a1a2* en Underhill (2014)) es más común (>30%) en la región de Altai, en el sur de Siberia, en Rusia, y aparece en Kirguistán (6%) y en todas las poblaciones iraníes (1-8%). [2]
El virus R-Z2125 se encuentra con mayor frecuencia en Kirguistán y entre los pastunes afganos (>40%). Con una frecuencia de >10%, también se observa en otros grupos étnicos afganos y en algunas poblaciones del Cáucaso e Irán. [2]
El R-M434 es un subclado del Z2125. Se detectó en 14 personas (de 3667 personas analizadas), todas en un área geográfica restringida desde Pakistán hasta Omán . Esto probablemente refleja un evento de mutación reciente en Pakistán. [3]
El R-M560 es muy raro y sólo se observó en cuatro muestras: dos hablantes de burushaski (norte de Pakistán), un hazara (Afganistán) y un azerbaiyano iraní. [2]
El R-M780 se presenta con alta frecuencia en el sur de Asia: India, Pakistán, Afganistán y el Himalaya. Turquía comparte sublinajes R1a (12,1%). [60] Los gitanos de Eslovaquia comparten el 3% de R1a. [61] El grupo también se presenta en >3% en algunas poblaciones iraníes y está presente en >30% en gitanos de Croacia y Hungría. [2]
Distribución geográfica de R1a1a
Distribución de R1a (violeta) y R1b (rojo)
Prehistórico
En la Europa mesolítica, R1a es característico de los cazadores-recolectores orientales (EHG). [62] Se descubrió que un EHG masculino de la cultura Veretye enterrado en Peschanitsa cerca del lago Lacha en el óblast de Arkhangelsk , Rusia, alrededor del 10 700 a. C. era portador del haplogrupo paterno R1a5-YP1301 y del haplogrupo materno U4a . [63] [64] [62] Se descubrió que un hombre, llamado PES001, de Peschanitsa en el noroeste de Rusia, portaba R1a5, y data de al menos 10 600 años atrás. [5] Más ejemplos incluyen a los hombres Minino II (V) y Minino II (I/1), el primero portando R1a1 y el segundo R1a respectivamente, el primero con 10 600 años y el segundo al menos 10 400 años respectivamente, ambos de Minino en el noroeste de Rusia. [65] Se ha descubierto que un varón mesolítico de Carelia entre el 8800 y el 7950 a. C. es portador del haplogrupo R1a. [66] Un varón mesolítico enterrado en Deriivka entre el 7000 y el 6700 a. C. era portador del haplogrupo paterno R1a y del materno U5a2a . [17] Otro varón de Carelia entre el 5500 y el 5000 a. C., considerado un EHG, era portador del haplogrupo R1a. [15] Se ha determinado que un varón de la cultura de la cerámica de peine en Kudruküla entre el 5900 y el 3800 a. C. es portador del R1a y del materno U2e1 . [67] Según el arqueólogo David Anthony, el R1a-Z93 paterno se encontró en el río Oskol cerca de un koljós "Alexandria" ya no existente, Ucrania, entre el 5000 y el 6000 a. C. 4000 a. C., "la muestra más antigua conocida que muestra la adaptación genética a la persistencia de la lactasa (13910-T)". [68] Se ha encontrado R1a en la cultura de la Cerámica Cordada , [69] [70] en la que es predominante. [71] Los varones examinados de la cultura Fatyanovo de la Edad del Bronce pertenecen enteramente a R1a, específicamente al subclado R1a-Z93. [62] [63] [72]
El haplogrupo R1a se ha encontrado posteriormente en fósiles antiguos asociados a la cultura de los campos de urnas ; [73] así como en el entierro de los restos de las culturas Sintashta , [16] Andronovo , [74] Pazyryk , [75] Tagar , [74] Tashtyk , [74] y Srubnaya , los habitantes del antiguo Tanais , [76] en las momias de Tarim , [77] y la aristocracia de Xiongnu . [78] Los restos esqueléticos de un padre y sus dos hijos, de un yacimiento arqueológico descubierto en 2005 cerca de Eulau (en Sajonia-Anhalt , Alemania ) y que data de alrededor de 2600 a. C., dieron positivo para el marcador Y-SNP SRY10831.2. El número de Ysearch para los restos de Eulau es 2C46S. El clado ancestral estuvo presente en Europa hace al menos 4600 años, en asociación con un sitio de la cultura de cerámica cordada muy extendida . [69]
Europa
En Europa, el subclado R1a1a es principalmente característico de las poblaciones baltoeslavas, con dos excepciones: los eslavos del sur y los rusos del norte. [79] La frecuencia más alta de R1a1a en Europa se observa en los sorbios (63%), [80] un grupo étnico eslavo occidental , seguido de los húngaros (60%). [12] Otros grupos con R1a1a significativo, que van desde el 27% hasta el 58%, incluyen a los checos , polacos , eslovenos , eslovacos , moldavos , bielorrusos , rusinos , ucranianos y rusos . [79] [80] [12] La frecuencia de R1a disminuye en las poblaciones del noreste de Rusia hasta el 20%–30%, en contraste con el centro-sur de Rusia, donde su frecuencia es el doble. En los países bálticos , las frecuencias de R1a1a disminuyen desde Lituania (45%) hasta Estonia (alrededor del 30%). [81] [82] [83] [12] [84]
También hay una presencia significativa en pueblos de ascendencia germánica , con niveles más altos en Noruega , Suecia e Islandia , donde entre el 20 y el 30% de los hombres están en R1a1a. [85] [86] Los vikingos y normandos también pueden haber llevado el linaje R1a1a más lejos, lo que representa al menos parte de la pequeña presencia en las Islas Británicas , las Islas Canarias y Sicilia . [87] [88] El haplogrupo R1a1a promedia entre el 20 y el 30% en alemanes , con un pico en Rostock del 31,3%. [89] R1a1a se encuentra con una frecuencia muy baja entre los holandeses (3,7%) [12] y está prácticamente ausente en los daneses . [90]
En el sur de Europa, el R1a1a no es común, pero se han encontrado niveles significativos en algunos lugares, como en el valle del Pas en el norte de España , áreas de Venecia y Calabria en Italia . [91] [ se necesita una mejor fuente ] Los Balcanes muestran una amplia variación entre áreas con niveles significativos de R1a1a, por ejemplo, 36-39% en Eslovenia , [92] 27-34% en Croacia , [82] [93] [94] [95] [96] y más del 30% en Macedonia griega , pero menos del 10% en Albania , Kosovo y partes de Grecia al sur de la garganta del Olimpo. [97] [83] [12]
R1a está virtualmente compuesto solo por el subclado Z284 en Escandinavia . En Eslovenia, el subclado principal es Z282 (Z280 y M458), aunque el subclado Z284 se encontró en una muestra de un esloveno. Hay una representación insignificante de Z93 en Turquía , 12,1% [60] [2] Los eslavos occidentales y húngaros se caracterizan por una alta frecuencia del subclado M458 y una baja Z92, un subclado de Z280. Cientos de muestras eslovenas y checas carecen del subclado Z92 de Z280, mientras que polacos, eslovacos, croatas y húngaros solo muestran una frecuencia muy baja de Z92. [2] Los bálticos , eslavos orientales , serbios , macedonios , búlgaros y rumanos demuestran una proporción Z280>M458 y una alta, hasta prevaleciente participación de Z92. [2] Los bálticos y los eslavos orientales tienen los mismos subclados y frecuencias similares en una filogenia más detallada de los subclados. [98] [99] El genetista ruso Oleg Balanovsky especuló que hay un predominio del sustrato preeslavo asimilado en la genética de las poblaciones eslavas orientales y occidentales, según él la estructura genética común que contrasta a los eslavos orientales y bálticos de otras poblaciones puede sugerir la explicación de que el sustrato preeslavo de los eslavos orientales y occidentales consistía más significativamente en hablantes del báltico, que en un momento precedieron a los eslavos en las culturas de la estepa euroasiática según referencias arqueológicas y toponímicas. [nota 16]
Asia
Asia central
Zerjal et al. (2002) encontraron R1a1a en el 64% de una muestra de tayikos de Tayikistán y en el 63% de una muestra de kirguisos de Kirguistán . [100]
Haber et al. (2012) encontraron R1a1a-M17 en el 26,0% (53/204) de un conjunto de muestras de Afganistán , incluido el 60% (3/5) de una muestra de nuristaníes , el 51,0% (25/49) de una muestra de pastunes, el 30,4% (17/56) de una muestra de tayikos, el 17,6% (3/17) de una muestra de uzbekos, el 6,7% (4/60) de una muestra de hazaras y en el único individuo turcomano muestreado. [101]
Di Cristofaro et al. (2013) encontraron R1a1a-M198/M17 en el 56,3% (49/87) de un par de muestras de pastunes de Afganistán (incluidos 20/34 o el 58,8% de una muestra de pastunes de Baghlan y 29/53 o el 54,7% de una muestra de pastunes de Kunduz ), 29,1% (37/127) de un grupo de muestras de uzbekos de Afganistán (incluidos 28/94 o el 29,8% de una muestra de uzbekos de Jawzjan , 8/28 o el 28,6% de una muestra de uzbekos de Sar-e Pol y 1/5 o el 20% de una muestra de uzbekos de Balkh ), 27,5% (39/142) de un grupo de muestras de tayikos de Afganistán (incluidos 22/5 ... 40,7% de una muestra de tayikos de Balkh , 9/35 o 25,7% de una muestra de tayikos de Takhar , 4/16 o 25,0% de una muestra de tayikos de Samangan , y 4/37 o 10,8% de una muestra de tayikos de Badakhshan ), 16,2% (12/74) de una muestra de turcomanos de Jawzjan , y 9,1% (7/77) de un par de muestras de hazara de Afganistán (incluyendo 7/69 o 10,1% de una muestra de hazara de Bamiyán y 0/8 o 0% de una muestra de hazara de Balkh ). [102]
Malyarchuk et al. (2013) encontraron R1a1-SRY10831.2 en el 30,0 % (12/40) de una muestra de tayikos de Tayikistán. [103]
Ashirbekov et al. (2017) encontraron R1a-M198 en el 6,03% (78/1294) de un conjunto de muestras de kazajos de Kazajstán . Se observó R1a-M198 con una frecuencia mayor que el promedio en las muestras del estudio de las siguientes tribus kazajas: 13/41 = 31,7 % de una muestra de Suan, 8/29 = 27,6 % de una muestra de Oshaqty, 6/30 = 20,0 % de una muestra de Qozha, 4/29 = 13,8% de una muestra de Qypshaq, 1/8 = 12,5% de una muestra de Tore, 9/86 = 10,5% de una muestra de Jetyru, 4/50 = 8,0% de una muestra de muestra de Argyn, 1/13 = 7,7% de una muestra de Shanyshqyly, 8/122 = 6,6% de una muestra de Alimuly, 3/46 = 6,5% de una muestra de Alban. También se observó R1a-M198 en 5/42 = 11,9% de una muestra de kazajos de afiliación tribal no declarada. [104]
Asia del Sur
En el sur de Asia, el R1a1a se ha observado a menudo en varios grupos demográficos. [34] [33]
Además de estos, los estudios muestran altos porcentajes en grupos regionalmente diversos, como los manipuris (50%) [3] en el extremo noreste y entre los punjabis (47%) [28] en el extremo noroeste.
A Chinese paper published in 2018 found R1a-Z94 in 38.5% (15/39) of a sample of Keriyalik Uyghurs from Darya Boyi / Darya Boye Village, Yutian County, Xinjiang (于田县达里雅布依乡), R1a-Z93 in 28.9% (22/76) of a sample of Dolan Uyghurs from Horiqol township, Awat County, Xinjiang (阿瓦提县乌鲁却勒镇), and R1a-Z93 in 6.3% (4/64) of a sample of Loplik Uyghurs from Karquga / Qarchugha Village, Yuli County, Xinjiang (尉犁县喀尔曲尕乡). R1a(xZ93) was observed only in one of 76 Dolan Uyghurs.[111] Note that Darya Boyi Village is located in a remote oasis formed by the Keriya River in the Taklamakan Desert. A 2011 Y-DNA study found Y-dna R1a1 in 10% of a sample of southern Hui people from Yunnan, 1.6% of a sample of Tibetan people from Tibet (Tibet Autonomous Region), 1.6% of a sample of Xibe people from Xinjiang, 3.2% of a sample of northern Hui from Ningxia, 9.4% of a sample of Hazak (Kazakhs) from Xinjiang, and rates of 24.0%, 22.2%, 35.2%, 29.2% in 4 different samples of Uyghurs from Xinjiang, 9.1% in a sample of Mongols from Inner Mongolia. A different subclade of R1 was also found in 1.5% of a sample of northern Hui from Ningxia.[112] in the same study there were no cases of R1a detected at all in 6 samples of Han Chinese in Yunnan, 1 sample of Han in Guangxi, 5 samples of Han in Guizhou, 2 samples of Han in Guangdong, 2 samples of Han in Fujian, 2 samples of Han in Zhejiang, 1 sample of Han in Shanghai, 1 samples of Han in Jiangxi, 2 samples of Han in Hunan, 1 sample of Han in Hubei, 2 samples of Han in Sichuan, 1 sample of Han in Chongqing, 3 samples of Han in Shandong, 5 samples of Han in Gansu, 3 samples of Han in Jilin and 2 samples of Han in Heilongjiang.[113] 40% of Salars, 45.2% of Tajiks of Xinjiang, 54.3% of Dongxiang, 60.6% of Tatars and 68.9% of Kyrgyz in Xinjiang in northwestern China tested in one sample had R1a1-M17. Bao'an (Bonan) had the most haplogroup diversity of 0.8946±0.0305 while the other ethnic minorities in northwestern China had a high haplogroup diversity like Central Asians, of 0.7602±0.0546.[114]
In Eastern Siberia, R1a1a is found among certain indigenous ethnic groups including Kamchatkans and Chukotkans, and peaking in Itel'man at 22%.[115]
Southeast Asia
Y-haplogroups R1a-M420 and R2-M479 are found in Ede (8.3% and 4.2%) and Giarai (3.7% and 3.7%) peoples in Vietnam. The Cham additionally have haplogroups R-M17 (13.6%) and R-M124 (3.4%).
R1a1a1b2a2a (R-Z2123) and R1a1 are found in Khmer peoples from Thailand (3.4%) and Cambodia (7.2%) respectively. Haplogroup R1a1a1b2a1b (R-Y6) is also found among Kuy peoples (5%).
According to Changmai et. al (2022), these haplogroup frequencies originate from South Asians, who left a cultural and genetic legacy in Southeast Asia since the first millennium CE.[116]
West Asia
R1a1a has been found in various forms, in most parts of Western Asia, in widely varying concentrations, from almost no presence in areas such as Jordan, to much higher levels in parts of Kuwait and Iran. The Shimar (Shammar) Bedouin tribe in Kuwait show the highest frequency in the Middle East at 43%.[117][118][119]
Wells 2001, noted that in the western part of the country, Iranians show low R1a1a levels, while males of eastern parts of Iran carried up to 35% R1a1a. Nasidze et al. 2004 found R1a1a in approximately 20% of Iranian males from the cities of Tehran and Isfahan. Regueiro 2006 in a study of Iran, noted much higher frequencies in the south than the north.
Di Cristofaro et al. (2013) found haplogroup R1a in 9.68% (18/186) of a set of samples from Iran, though with a large variance ranging from 0% (0/18) in a sample of Iranians from Tehran to 25% (5/20) in a sample of Iranians from Khorasan and 27% (3/11) in a sample of Iranians of unknown provenance. All Iranian R1a individuals carried the M198 and M17 mutations except one individual in a sample of Iranians from Gilan (n=27), who was reported to belong to R1a-SRY1532.2(xM198, M17).[102]
Malyarchuk et al. (2013) found R1a1-SRY10831.2 in 20.8% (16/77) of a sample of Persians collected in the provinces of Khorasan and Kerman in eastern Iran, but they did not find any member of this haplogroup in a sample of 25 Kurds collected in the province of Kermanshah in western Iran.[103]
Further to the north of these Western Asian regions on the other hand, R1a1a levels start to increase in the Caucasus, once again in an uneven way. Several populations studied have shown no sign of R1a1a, while highest levels so far discovered in the region appears to belong to speakers of the Karachay-Balkar language among whom about one quarter of men tested so far are in haplogroup R1a1a.[3]
Historic naming of R1a
The historic naming system commonly used for R1a was inconsistent in different published sources, because it changed often; this requires some explanation.
In 2002, the Y Chromosome Consortium (YCC) proposed a new naming system for haplogroups (YCC 2002), which has now become standard. In this system, names with the format "R1" and "R1a" are "phylogenetic" names, aimed at marking positions in a family tree. Names of SNP mutations can also be used to name clades or haplogroups. For example, as M173 is currently the defining mutation of R1, R1 is also R-M173, a "mutational" clade name. When a new branching in a tree is discovered, some phylogenetic names will change, but by definition all mutational names will remain the same.
The widely occurring haplogroup defined by mutation M17 was known by various names, such as "Eu19", as used in (Semino et al. 2000) in the older naming systems. The 2002 YCC proposal assigned the name R1a to the haplogroup defined by mutation SRY1532.2. This included Eu19 (i.e. R-M17) as a subclade, so Eu19 was named R1a1. Note, SRY1532.2 is also known as SRY10831.2[citation needed] The discovery of M420 in 2009 has caused a reassignment of these phylogenetic names.(Underhill et al. 2009 and ISOGG 2012) R1a is now defined by the M420 mutation: in this updated tree, the subclade defined by SRY1532.2 has moved from R1a to R1a1, and Eu19 (R-M17) from R1a1 to R1a1a.
More recent updates recorded at the ISOGG reference webpage involve branches of R-M17, including one major branch, R-M417.
^According to Family Tree,[who?] they diversified c. 5,000 years ago.[9]
^See also: "'Heavily sex-biased' population dispersals into the Indian Subcontinent (Silva et al. 2017)". Eurogenes Blog. March 28, 2017.[self-published source?]
^Semenov & Bulat (2016) refer to the following publications:
Haak, Wolfgang (2015). "Massive migration from the steppe is a source for Indo-European languages in Europe". Nature. 522 (7555): 207–211. arXiv:1502.02783. Bibcode:2015Natur.522..207H. bioRxiv10.1101/013433. doi:10.1038/NATURE14317. PMC 5048219. PMID 25731166.
Mathieson, Iain (2015). "Eight thousand years of natural selection in Europe". bioRxiv10.1101/016477.
Chekunova Е.М., Yartseva N.V., Chekunov М.К., Мazurkevich А.N. The First Results of the Genotyping of the Aboriginals and Human Bone Remains of the Archeological Memorials of the Upper Podvin'e. // Archeology of the lake settlements of IV—II Thousands BC: The chronology of cultures and natural environment and climatic rhythms. Proceedings of the International Conference, Devoted to the 50-year Research of the Pile Settlements on the North-West of Russia. St. Petersburg, November 13–15, 2014.
^Kivisild et al. (2003): "Haplogroup R1a, previously associated with the putative Indo-Aryan invasion, was found at its highest frequency in Punjab but also at a relatively high frequency (26%) in the Chenchu tribe. This finding, together with the higher R1a-associated short tandem repeat diversity in India and Iran compared with Europe and central Asia, suggests that southern and western Asia might be the source of this haplogroup."[28]
^Sengupta (2006): "We found that the influence of Central Asia on the pre-existing gene pool was minor. The ages of accumulated microsatellite variation in the majority of Indian haplogroups exceed 10,000–15,000 years, which attests to the antiquity of regional differentiation. Therefore, our data do not support models that invoke a pronounced recent genetic input from Central Asia to explain the observed genetic variation in South Asia."
^South-Asian origins: * Sahoo et al. (2006): "... one should expect to observe dramatically lower genetic variation among Indian Rla lineages. In fact, the opposite is true: the STR haplotype diversity on the background of R1a in Central Asia (and also in Eastern Europe) has already been shown to be lower than that in India (6). Rather, the high incidence of R1* and Rla throughout Central Asian European populations (without R2 and R* in most cases) is more parsimoniously explained by gene flow in the opposite direction, possibly with an early founder effect in South or West Asia.[36] * Sharma et al. (2009): "A peculiar observation of the highest frequency (up to 72.22%) of Y-haplogroup R1a1* in Brahmins hinted at its presence as a founder lineage for this caste group. Further, observation of R1a1* in different tribal population groups, existence of Y-haplogroup R1a* in ancestors and extended phylogenetic analyses of the pooled dataset of 530 Indians, 224 Pakistanis and 276 Central Asians and Eurasians bearing the R1a1* haplogroup supported the autochthonous origin of R1a1 lineage in India and a tribal link to Indian Brahmins. However, it is important to discover novel Y-chromosomal binary marker(s) for a higher resolution of R1a1* and confirm the present conclusions."
^Though Sengupta (2006) did concede that "[R1a1 and R2] could have actually arrived in southern India from a southwestern Asian source region multiple times." In full: "The widespread geographic distribution of HG R1a1-M17 across Eurasia and the current absence of informative subdivisions defined by binary markers leave uncertain the geographic origin of HG R1a1-M17. However, the contour map of R1a1-M17 variance shows the highest variance in the northwestern region of India ... The question remains of how distinctive is the history of L1 relative to some or all of R1a1 and R2 representatives. This uncertainty neutralizes previous conclusions that the intrusion of HGs R1a1 and R2 from the northwest in Dravidian-speaking southern tribes is attributable to a single recent event. [R1a1 and R2] could have actually arrived in southern India from a southwestern Asian source region multiple times, with some episodes considerably earlier than others. Considerable archeological evidence exists regarding the presence of Mesolithic peoples in India (Kennedy 2000), some of whom could have entered the subcontinent from the northwest during the late Pleistocene epoch. The high variance of R1a1 in India (table 12), the spatial frequency distribution of R1a1 microsatellite variance clines (fig. 4), and expansion time (table 11) support this view."[33]
^Lalueza-Fox: "Some years ago, local scientists supported the view that the existence of an R1a Y chromosome was not attributable to a foreign gene flow but instead that this lineage had emerged on the subcontinent and spread from there. But the phylogenetic reconstruction of this haplogroup did not support this view."[37]
^Yet, Haak et al. also explicitly state: "a type of Near Eastern ancestry different from that which was introduced by early farmers".[clarification needed][39]
^According to Family Tree DNA, L664 formed 4,700 ybp, that is, 2,700 BCE.[9]
^Lazaridis, Twitter, 18 June 2016: "I1635 (Armenia_EBA) is R1b1-M415(xM269). We'll be sure to include in the revision. Thanks to the person who noticed! #ILovePreprints."[unreliable source?] See also "Big deal of 2016: the territory of present-day Iran cannot be the Indo-European homeland". Eurogenes Blog. November 26, 2016,[unreliable source?] for a discussion of the same topic.
^See map for M780 distribution at Dieneke's Anthropology Blog, Major new article on the deep origins of Y-haplogroup R1a (Underhill et al. 2014)[43]
^According to Family Tree DNA, M780 formed 4700 ybp.[9] This dating coincides with the eastward movement between 2800 and 2600 BCE of the Yamnaya culture into the region of the Poltavka culture, a predecessor of the Sintashta culture, from which the Indo-Iranians originated. M780 is concentrated in the Ganges Valley, the locus of the classic Vedic society.
^Poznik et al. (2016) calculate with a generation time of 30 years; a generation time of 20 years yields other results.
^"The evidence that the Steppe_MLBA [Middle to Late Bronze Age] cluster is a plausible source for the Steppe ancestry in South Asia is also supported by Y chromosome evidence, as haplogroup R1a which is of the Z93 subtype common in South Asia today [Underhill et al. (2014), Silva et al. (2017)] was of high frequency in Steppe_MLBA (68%) (16), but rare in Steppe_EMBA [Early to Middle Bronze Age] (absent in our data)."[44]
^Балановский (2015), p. 208 (in Russian)Прежде всего, это преобладание в славянских популяциях дославянского субстрата — двух ассимилированных ими генетических компонентов – восточноевропейского для западных и восточных славян и южноевропейского для южных славян...Можно с осторожностью предположить, что ассимилированный субстратмог быть представлен по преимуществу балтоязычными популяциями. Действительно, археологические данные указыва ют на очень широкое распространение балтских групп перед началом расселения славян. Балтскийсубстрату славян (правда, наряду с финно-угорским) выявляли и антропологи. Полученные нами генетические данные — и на графиках генетических взаимоотношений, и по доле общих фрагментов генома — указывают, что современные балтские народы являются ближайшими генетически ми соседями восточных славян. При этом балты являются и лингвистически ближайшими род ственниками славян. И можно полагать, что к моменту ассимиляции их генофонд не так сильно отличался от генофонда начавших свое широкое расселение славян. Поэтому если предположить,что расселяющиеся на восток славяне ассимилировали по преимуществу балтов, это может объяснить и сходство современных славянских и балтских народов друг с другом, и их отличия от окружающих их не балто-славянских групп Европы...В работе высказывается осторожное предположение, что ассимилированный субстрат мог быть представлен по преимуществу балтоязычными популяциями. Действительно, археологические данные указывают на очень широкое распространение балтских групп перед началом расселения славян. Балтский субстрат у славян (правда, наряду с финно-угорским) выявляли и антропологи. Полученные в этой работе генетические данные — и на графиках генетических взаимоотношений, и по доле общих фрагментов генома — указывают, что современные балтские народы являются ближайшими генетическими соседями восточных славян.
References
^ a b c d e fSharma et al. 2009.
^ a b c d e f g h i j k l m n o p q r s t uUnderhill et al. 2014.
^ a b c d e f g h i j k l m n o p q rUnderhill et al. 2009.
^ a bUnderhill et al. 2014, p. 130.
^ a bSaag, Lehti; Vasilyev, Sergey V.; Varul, Liivi; Kosorukova, Natalia V.; Gerasimov, Dmitri V.; Oshibkina, Svetlana V.; Griffith, Samuel J.; Solnik, Anu; Saag, Lauri; D'Atanasio, Eugenia; Metspalu, Ene (January 2021). "Genetic ancestry changes in Stone to Bronze Age transition in the East European plain". Science Advances. 7 (4): eabd6535. Bibcode:2021SciA....7.6535S. doi:10.1126/sciadv.abd6535. PMC 7817100. PMID 33523926.
^Haak, Wolfgang; Lazaridis, Iosif; Patterson, Nick; Rohland, Nadin; Mallick, Swapan; Llamas, Bastien; Brandt, Guido; Nordenfelt, Susanne; Harney, Eadaoin; Stewardson, Kristin; Fu, Qiaomei (February 10, 2015). "Massive migration from the steppe is a source for Indo-European languages in Europe". bioRxiv: 013433. arXiv:1502.02783. doi:10.1101/013433. S2CID 196643946. Archived from the original on December 23, 2019. Retrieved February 8, 2021.
^Narasimhan, Vagheesh M.; Patterson, Nick; Moorjani, Priya; Rohland, Nadin; Bernardos, Rebecca; Mallick, Swapan; Lazaridis, Iosif; Nakatsuka, Nathan; Olalde, Iñigo; Lipson, Mark; Kim, Alexander M. (September 6, 2019). "The formation of human populations in South and Central Asia". Science. 365 (6457): eaat7487. doi:10.1126/science.aat7487. PMC 6822619. PMID 31488661. Y chromosome haplogroup types R1b or R1a not represented in Iran and Turan in this period ...
^ a b c d e f g h i j"R1a tree". YFull. Archived from the original on August 19, 2016. Retrieved July 15, 2016.
^ a b cMirabal et al. 2009.
^Zerjal, T.; et al. (1999). "The use of Y-chromosomal DNA variation to investigate population history: recent male spread in Asia and Europe". In Papiha, S. S.; Deka, R. & Chakraborty, R. (eds.). Genomic diversity: applications in human population genetics. New York: Kluwer Academic/Plenum Publishers. pp. 91–101. ISBN 978-0-3064-6295-5.
^ a b c d e fSemino et al. 2000.
^ a bWells 2001.
^ a b cPamjav et al. 2012.
^ a bHaak et al. 2015.
^ a bAllentoft et al. 2015.
^ a bMathieson et al. 2015.
^ a b cSilva et al. 2017.
^Joseph, Tony (June 16, 2017). "How genetics is settling the Aryan migration debate". The Hindu. Archived from the original on October 4, 2023. Retrieved June 2, 2019.
^Anthony 2007.
^Anthony & Ringe 2015.
^ a bHaak et al. 2015, p. 5.
^Semenov & Bulat 2016.
^Haber et al. 2012"R1a1a7-M458 was absent in Afghanistan, suggesting that R1a1a-M17 does not support, as previously thought [47], expansions from the Pontic Steppe [3], bringing the Indo-European languages to Central Asia and India."
^Klejn, Leo S. (April 22, 2017). "The Steppe Hypothesis of Indo-European Origins Remains to be Proven". Acta Archaeologica. 88 (1): 193–204. doi:10.1111/j.1600-0390.2017.12184.x. ISSN 0065-101X. Archived from the original on December 25, 2022. Retrieved November 23, 2022. "As for the Y-chromosome, it was already noted in Haak, Lazaridis et al. (2015) that the Yamnaya from Samara had Y-chromosomes which belonged to R-M269 but did not belong to the clade common in Western Europe (p. 46 of supplement). Also, not a single R1a in Yamnaya unlike Corded Ware (R1a-dominated)."
^Koch, John T.; Cunliffe, Barry (2016). Celtic from the West 3: Atlantic Europe in the Metal Ages. Oxbow Books. p. 634. ISBN 978-1-78570-228-0. Archived from the original on November 23, 2022. Retrieved November 23, 2022.
^Semenov & Bulat 2016, p. 41.
^ a b c dKivisild et al. 2003.
^Sengupta S, Zhivotovsky LA, King R, Mehdi SQ, Edmonds CA, Chow CE, et al. (February 2006). "Polarity and temporality of high-resolution y-chromosome distributions in India identify both indigenous and exogenous expansions and reveal minor genetic influence of Central Asian pastoralists". American Journal of Human Genetics. 78 (2): 202–221. doi:10.1086/499411. PMC 1380230. PMID 16400607."Although considerable cultural impact on social hierarchy and language in South Asia is attributable to the arrival of nomadic Central Asian pastoralists, genetic data (mitochondrial and Y chromosomal) have yielded dramatically conflicting inferences on the genetic origins of tribes and castes of South Asia. We sought to resolve this conflict, using high-resolution data on 69 informative Y-chromosome binary markers and 10 microsatellite markers from a large set of geographically, socially, and linguistically representative ethnic groups of South Asia. We found that the influence of Central Asia on the pre-existing gene pool was minor. The ages of accumulated microsatellite variation in the majority of Indian haplogroups exceed 10,000–15,000 years, which attests to the antiquity of regional differentiation. Therefore, our data do not support models that invoke a pronounced recent genetic input from Central Asia to explain the observed genetic variation in South Asia. R1a1 and R2 haplogroups indicate demographic complexity that is inconsistent with a recent single history.ASSOCIATED MICROSATELLITE ANALYSES OF THE HIGH-FREQUENCY R1A1 HAPLOGROUP CHROMOSOMES INDICATE INDEPENDENT RECENT HISTORIES OF THE INDUS VALLEY AND THE PENINSULAR INDIAN REGION."
^Thanseem I, Thangaraj K, Chaubey G, Singh VK, Bhaskar LV, Reddy BM, et al. (August 2006). "Genetic affinities among the lower castes and tribal groups of India: inference from Y chromosome and mitochondrial DNA". BMC Genetics. 7: 42. doi:10.1186/1471-2156-7-42. PMC 1569435. PMID 16893451.
^Sahoo S, Singh A, Himabindu G, Banerjee J, Sitalaximi T, Gaikwad S, et al. (January 2006). "A prehistory of Indian Y chromosomes: evaluating demic diffusion scenarios". Proceedings of the National Academy of Sciences of the United States of America. 103 (4): 843–848. Bibcode:2006PNAS..103..843S. doi:10.1073/pnas.0507714103. PMC 1347984. PMID 16415161.
^Thangaraj K, Naidu BP, Crivellaro F, Tamang R, Upadhyay S, Sharma VK, et al. (December 2010). Cordaux R (ed.). "The influence of natural barriers in shaping the genetic structure of Maharashtra populations". PLOS ONE. 5 (12): e15283. Bibcode:2010PLoSO...515283T. doi:10.1371/journal.pone.0015283. PMC 3004917. PMID 21187967.
^ a b c d e fSengupta 2006.
^ a b c d eSahoo et al. 2006.
^ a bThangaraj et al. 2010.
^Sahoo et al. 2006, p. 845-846.
^ a bLalueza-Fox, C. (2022). Inequality: A Genetic History. MIT Press. pp. 81–82. ISBN 978-0-262-04678-7. Archived from the original on July 16, 2023. Retrieved July 16, 2023.
^Narasimhan et al. 2019.
^Haak et al. 2015, p. 4.
^ a b cMascarenhas et al. 2015, p. 9.
^ a bPoznik et al. 2016, p. 5.
^Arame's English blog, Y DNA from ancient Near East Archived November 27, 2016, at the Wayback Machine
^"Dienekes' Anthropology Blog: Major new article on the deep origins of Y-haplogroup R1a (Underhill et al. 2014)". March 27, 2014. Archived from the original on December 20, 2019. Retrieved December 20, 2019.[unreliable source?]
^ a bNarasimhan et al. 2018.
^ a b c"About Us". Family Tree DNA. Archived from the original on August 15, 2019. Retrieved December 20, 2019.
^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa"ISOGG 2017 Y-DNA Haplogroup R". isogg.org. Archived from the original on February 10, 2007. Retrieved December 20, 2019.
^ a b c d e f g h i j k"Haplogroup R (Y-DNA) - SNPedia". www.snpedia.com. Archived from the original on May 5, 2018. Retrieved December 20, 2019.
^Karafet et al. 2014.
^ a b c d e f g h i j k l mUnderhill et al. 2014, p. 125.
^"R1a in Yamnaya". Eurogenes Blog. March 21, 2016. Archived from the original on May 5, 2018. Retrieved December 20, 2019.
^"Y-DNA Haplogroup R and its Subclades". International Society of Genetic Genealogy (ISOGG). Archived from the original on March 30, 2019. Retrieved January 8, 2011.
^Krahn, Thomas. "Draft Y-Chromosome Tree". Family Tree DNA. Archived from the original on May 26, 2013. Retrieved December 7, 2012.
^Regueiro 2006.
^Freder, Janine (2010). Die mittelalterlichen Skelette von Usedom: Anthropologische Bearbeitung unter besonderer Berücksichtigung des ethnischen Hintergrundes [Anthropological investigation in due consideration of the ethnical background] (Thesis) (in German). Freie Universität Berlin. p. 86. doi:10.17169/refubium-8995.
^https://cyberleninka.ru/article/n/tyurki-kavkaza-sravnitelnyy-analiz-genofondov-po-dannym-o-y-hromosome Archived November 7, 2023, at the Wayback Machine "высокая частота R1a среди кубанских ногайцев (субветвь R1a1a1g-M458 забирает 18%"
^Underhill, P. A.; et al. (2009). "Separating the post-Glacial coancestry of European and Asian y chromosomes within haplogroup R1a". European Journal of Human Genetics. 18 (4): 479–484. doi:10.1038/ejhg.2009.194. PMC 2987245. PMID 19888303.
^Gwozdz, Peter (August 6, 2018). "Polish Y-DNA Clades". Archived from the original on July 15, 2016. Retrieved July 15, 2016.
^Pawlowski et al. 2002.
^ a bGwozdz 2009.
^ a bKars, M. E.; Başak, A. N.; Onat, O. E.; Bilguvar, K.; Choi, J.; Itan, Y.; Çağlar, C.; Palvadeau, R.; Casanova, J. L.; Cooper, D. N.; Stenson, P. D.; Yavuz, A.; Buluş, H.; Günel, M.; Friedman, J. M.; Özçelik, T. (2021). "The genetic structure of the Turkish population reveals high levels of variation and admixture". Proceedings of the National Academy of Sciences of the United States of America. 118 (36): e2026076118. Bibcode:2021PNAS..11826076K. doi:10.1073/pnas.2026076118. PMC 8433500. PMID 34426522.
^Saag et al. 2020, Supplementary Data 2, Rows 5-49.
^Schweitzer, D. (March 23, 2008). "Lichtenstein Cave Data Analysis" (PDF). dirkschweitzer.net. Archived from the original (PDF) on August 14, 2011. Summary in English of Schilz (2006).
^ a b cKeyser et al. 2009.
^Ricaut et al. 2004.
^Korniyenko, I. V.; Vodolazhsky D. I. "Использование нерекомбинантных маркеров Y-хромосомы в исследованиях древних популяций (на примере поселения Танаис)" [The use of non-recombinant markers of the Y-chromosome in the study of ancient populations (on the example of the settlement of Tanais)]. Материалы Донских антропологических чтений [Materials of the Don Anthropological Readings]. Rostov-on-Don: Rostov Research Institute of Oncology, 2013.
^Chunxiang Li et al. 2010.
^Kim et al. 2010.
^ a bBalanovsky et al. 2008.
^ a bBehar et al. 2003.
^Kasperaviciūte, Kucinskas & Stoneking 2005.
^ a bBattaglia et al. 2008.
^ a bRosser et al. 2000.
^Tambets et al. 2004.
^Bowden et al. 2008.
^Dupuy et al. 2005.
^Passarino et al. 2002.
^Capelli et al. 2003.
^Kayser et al. 2005.
^Sanchez, J; Børsting, C; Hallenberg, C; Buchard, A; Hernandez, A; Morling, N (2003). "Multiplex PCR and minisequencing of SNPs—a model with 35 Y chromosome SNPs". Forensic Science International. 137 (1): 74–84. doi:10.1016/S0379-0738(03)00299-8. PMID 14550618.
^Scozzari et al. 2001.
^Underhill, Peter A. (January 1, 2015). "The phylogenetic and geographic structure of Y-chromosome haplogroup R1a". European Journal of Human Genetics. 23 (1): 124–131. doi:10.1038/ejhg.2014.50. PMC 4266736. PMID 24667786.
^L. Barać; et al. (2003). "Y chromosomal heritage of Croatian population and its island isolates" (PDF). European Journal of Human Genetics. 11 (7): 535–42. doi:10.1038/sj.ejhg.5200992. PMID 12825075. S2CID 15822710. Archived from the original (PDF) on December 17, 2012. Retrieved September 10, 2009.
^S. Rootsi; et al. (2004). "Phylogeography of Y-Chromosome Haplogroup I Reveals Distinct Domains of Prehistoric Gene Flow in Europe" (PDF). American Journal of Human Genetics. 75 (1): 128–137. doi:10.1086/422196. PMC 1181996. PMID 15162323. Archived from the original (PDF) on September 5, 2020. Retrieved February 13, 2021.
^M. Peričić; et al. (2005). "High-resolution phylogenetic analysis of southeastern Europe traces major episodes of paternal gene flow among Slavic populations". Molecular Biology and Evolution. 22 (10): 1964–75. doi:10.1093/molbev/msi185. PMID 15944443.
^M. Peričić; et al. (2005). "Review of Croatian Genetic Heritage as Revealed by Mitochondrial DNA and Y Chromosomal Lineages". Croatian Medical Journal. 46 (4): 502–513. PMID 16100752.
^Pericić et al. 2005.
^"Untitled". pereformat.ru (in Russian). Archived from the original on March 15, 2016. Retrieved May 29, 2017.
^"Untitled". www.rodstvo.ru. Archived from the original on September 16, 2021. Retrieved May 29, 2017.
^Zerjal et al. 2002.
^Haber et al. 2012.
^ a bDi Cristofaro et al. 2013.
^ a bMalyarchuk et al. 2013.
^Ashirbekov et al. 2017.
^Shah 2011.
^Arunkumar 2012.
^Toomas Kivisild; Siiri Rootsi; Mait Metspalu; Ene Metspalu; Juri Parik; Katrin Kaldma; Esien Usanga; Sarabjit Mastana; Surinder S. Papiha; Richard Villems. "The Genetics of Language and Farming Spread in India" (PDF). In P. Bellwwood; C. Renfrew (eds.). Examining the farming/language dispersal hypothesis. McDonald Institute Monographs. Cambridge University. pp. 215–222. Archived from the original (PDF) on February 19, 2006. Retrieved December 20, 2019.
^Fornarino et al. 2009.
^Wang et al. 2003.
^Zhou et al. 2007.
^Liu Shu-hu et al. 2018.
^Zhong et al. 2011.
^Zhong, Hua; Shi, Hong; Qi, Xue-Bin; Duan, Zi-Yuan; Tan, Ping-Ping; Jin, Li; Su, Bing; Ma, Runlin Z. (2011). "Extended Y Chromosome Investigation Suggests Postglacial Migrations of Modern Humans into East Asia via the Northern Route". Molecular Biology and Evolution. 28 (1): 717–727. doi:10.1093/molbev/msq247. PMID 20837606.
^Shou, Wei-Hua; Qiao, Wn-Fa; Wei, Chuan-Yu; Dong, Yong-Li; Tan, Si-Jie; Shi, Hong; Tang, Wen-Ru; Xiao, Chun-Jie (2010). "Y-chromosome distributions among populations in Northwest China identify significant contribution from Central Asian pastoralists and lesser influence of western Eurasians". Journal of Human Genetics. 55 (5): 314–322. doi:10.1038/jhg.2010.30. PMID 20414255. S2CID 23002493.
^Lell et al. 2002.
^Changmai, Piya; Jaisamut, Kitipong; Kampuansai, Jatupol; et al. (2022). "Indian genetic heritage in Southeast Asian populations". PLOS Genetics. 18 (2): e1010036. doi:10.1371/journal.pgen.1010036. PMC 8853555. PMID 35176016.
^Mohammad et al. 2009.
^Nasidze et al. 2004.
^Nasidze et al. 2005.
^Grugni et al. 2012.
Sources
Allentoft, Morten E.; Sikora, Martin; Sjögren, Karl-Göran; Rasmussen, Simon; Rasmussen, Morten; Stenderup, Jesper; Damgaard, Peter B.; Schroeder, Hannes; et al. (2015). "Population genomics of Bronze Age Eurasia". Nature. 522 (7555): 167–172. Bibcode:2015Natur.522..167A. doi:10.1038/nature14507. PMID 26062507. S2CID 4399103. Archived from the original on July 13, 2019. Retrieved July 23, 2019.
Anthony, David W. (2007), The Horse The Wheel And Language. How Bronze-Age Riders From the Eurasian Steppes Shaped The Modern World, Princeton University Press
Anthony, David (Spring–Summer 2019). "Archaeology, Genetics, and Language in the Steppes: A Comment on Bomhard". Journal of Indo-European Studies. 47 (1–2). Archived from the original on May 3, 2024. Retrieved January 9, 2020.
Anthony, David; Ringe, Don (2015), "The Indo-European Homeland from Linguistic and Archaeological Perspectives", Annual Review of Linguistics, 1: 199–219, doi:10.1146/annurev-linguist-030514-124812
Shah, A. M.; Tamang, R.; Moorjani, P.; Rani, D. S.; Govindaraj, P.; Kulkarni, G.; Bhattacharya, T.; Mustak, M. S.; Bhaskar, L. V. K. S.; Reddy, A. G.; Gadhvi, D.; Gai, P. B.; Chaubey, G.; Patterson, N.; Reich, D.; Tyler-Smith, C.; Singh, L.; Thangaraj, K. (2011). "Indian Siddis: African Descendants with Indian Admixture". The American Journal of Human Genetics. 89 (1): 154–61. doi:10.1016/j.ajhg.2011.05.030. PMC 3135801. PMID 21741027.
ArunKumar, G; Soria-Hernanz, DF; Kavitha, VJ; Arun, VS; Syama, A; Ashokan, KS (2012). "Population Differentiation of Southern Indian Male Lineages Correlates with Agricultural Expansions Predating the Caste System". PLOS ONE. 7 (11): e50269. Bibcode:2012PLoSO...750269A. doi:10.1371/journal.pone.0050269. PMC 3508930. PMID 23209694.
Ashirbekov, E. E.; et al. (2017). "Distribution of Y-Chromosome Haplogroups of the Kazakh from the South Kazakhstan, Zhambyl, and Almaty Regions" (PDF). Reports of the National Academy of Sciences of the Republic of Kazakhstan. 6 (316): 85–95. Archived from the original (PDF) on September 23, 2021. Retrieved June 4, 2020.
Balanovsky O, Rootsi S, Pshenichnov A, Kivisild T, Churnosov M, Evseeva I, Pocheshkhova E, Boldyreva M, et al. (2008). "Two Sources of the Russian Patrilineal Heritage in Their Eurasian Context". American Journal of Human Genetics. 82 (1): 236–250. doi:10.1016/j.ajhg.2007.09.019. PMC 2253976. PMID 18179905.
Балановский, О. П. (November 30, 2015). Генофонд Европы (in Russian). KMK Scientific Press. ISBN 9785990715707. Archived from the original on May 3, 2024. Retrieved August 8, 2020.
Battaglia V, Fornarino S, Al-Zahery N, Olivieri A, Pala M, Myres NM, King RJ, Rootsi S, et al. (2008). "Y-chromosomal evidence of the cultural diffusion of agriculture in southeast Europe". European Journal of Human Genetics. 17 (6): 820–30. doi:10.1038/ejhg.2008.249. PMC 2947100. PMID 19107149.
Behar D, Thomas MG, Skorecki K, Hammer MF, Bulygina E, Rosengarten D, Jones AL, Held K, et al. (2003). "Multiple Origins of Ashkenazi Levites: Y Chromosome Evidence for Both Near Eastern and European Ancestries" (PDF). American Journal of Human Genetics. 73 (4): 768–779. doi:10.1086/378506. PMC 1180600. PMID 13680527. Archived (PDF) from the original on April 17, 2007. Retrieved April 23, 2007.
Bowden GR, Balaresque P, King TE, Hansen Z, Lee AC, Pergl-Wilson G, Hurley E, Roberts SJ, et al. (2008). "Excavating Past Population Structures by Surname-Based Sampling: The Genetic Legacy of the Vikings in Northwest England". Molecular Biology and Evolution. 25 (2): 301–309. doi:10.1093/molbev/msm255. PMC 2628767. PMID 18032405.
Brandit, G.; et al. (The Genographic Consortium) (2013). "Ancient DNA Reveals Key Stages in the Formation of Central European Mitochondrial Genetic Diversity". Science. 342 (6155): 257–261. Bibcode:2013Sci...342..257B. doi:10.1126/science.1241844. PMC 4039305. PMID 24115443.
Capelli C, Redhead N, Abernethy JK, Gratrix F, Wilson JF, Moen T, Hervig T, Richards M, et al. (2003). "A Y Chromosome Census of the British Isles" (PDF). Current Biology. 13 (11): 979–84. Bibcode:2003CBio...13..979C. doi:10.1016/S0960-9822(03)00373-7. PMID 12781138. S2CID 526263. Archived (PDF) from the original on July 8, 2021. Retrieved August 8, 2020. also at "University College London" (PDF). Archived (PDF) from the original on January 17, 2017. Retrieved January 29, 2007.
Chunxiang Li; Hongjie Li; Yinqiu Cui; Chengzhi Xie; Dawei Cai; Wenying Li; Victor H Mair; Zhi Xu; et al. (2010). "Evidence that a West-East admixed population lived in the Tarim Basin as early as the early Bronze Age". BMC Biology. 8 (1): 15. doi:10.1186/1741-7007-8-15. PMC 2838831. PMID 20163704.
Di Cristofaro J, Pennarun E, Mazières S, Myres NM, Lin AA, Temori SA, Metspalu M, Metspalu E, et al. (2013). "Afghan Hindu Kush: Where Eurasian Sub-Continent Gene Flows Converge". PLOS ONE. 8 (10). e76748. Bibcode:2013PLoSO...876748D. doi:10.1371/journal.pone.0076748. PMC 3799995. PMID 24204668.
Dupuy BM, Stenersen M, Lu TT, Olaisen B (2005). "Geographical heterogeneity of Y-chromosomal lineages in Norway" (PDF). Forensic Science International. 164 (1): 10–19. doi:10.1016/j.forsciint.2005.11.009. PMID 16337760. Archived (PDF) from the original on July 22, 2012. Retrieved November 18, 2009.
Fornarino, Simona; Pala, Maria; Battaglia, Vincenza; Maranta, Ramona; Achilli, Alessandro; Modiano, Guido; Torroni, Antonio; Semino, Ornella; et al. (2009). "Mitochondrial and Y-chromosome diversity of the Tharus (Nepal): a reservoir of genetic variation". BMC Evolutionary Biology. 9 (1): 154. Bibcode:2009BMCEE...9..154F. doi:10.1186/1471-2148-9-154. PMC 2720951. PMID 19573232.
Fu, Qiaomei; et al. (May 2, 2016). "The genetic history of Ice Age Europe". Nature. 534 (7606): 200–205. Bibcode:2016Natur.534..200F. doi:10.1038/nature17993. hdl:10211.3/198594. PMC 4943878. PMID 27135931.
Grugni V, Battaglia V, Kashani BH, Parolo S, Al-Zahery N, Achilli A, Olivieri A, Gandini F, Houshmand M, Sanati MH, Torroni A, Semino O (2012). "Ancient Migratory Events in the Middle East: New Clues from the Y-Chromosome Variation of Modern Iranians". PLOS ONE. 7 (7). e41252. Bibcode:2012PLoSO...741252G. doi:10.1371/journal.pone.0041252. PMC 3399854. PMID 22815981.
Gwozdz (2009). "Y-STR Mountains in Haplospace, Part II: Application to Common Polish Clades" (PDF). Journal of Genetic Genealogy. 5 (2). Archived (PDF) from the original on July 21, 2011. Retrieved November 28, 2009.
Haak, W.; Brandt, G.; Jong, H. N. d.; Meyer, C.; Ganslmeier, R.; Heyd, V.; Hawkesworth, C.; Pike, A. W. G.; et al. (2008). "Ancient DNA, Strontium isotopes, and osteological analyses shed light on social and kinship organization of the Later Stone Age". Proceedings of the National Academy of Sciences. 105 (47): 18226–18231. Bibcode:2008PNAS..10518226H. doi:10.1073/pnas.0807592105. PMC 2587582. PMID 19015520.
Haak, Wolfgang; Lazaridis, Iosif; Patterson, Nick; Rohland, Nadin; Mallick, Swapan; Llamas, Bastien; Brandt, Guido; Nordenfelt, Susanne; et al. (2015). "Massive migration from the steppe is a source for Indo-European languages in Europe". Nature. 522 (7555): 207–211. arXiv:1502.02783. Bibcode:2015Natur.522..207H. bioRxiv10.1101/013433. doi:10.1038/NATURE14317. PMC 5048219. PMID 25731166.
Haber M, Platt DE, Ashrafian Bonab M, Youhanna SC, Soria-Hernanz DF, Martínez-Cruz B, Douaihy B, Ghassibe-Sabbagh M, et al. (2012). "Afghanistan's ethnic groups share a Y-chromosomal heritage structured by historical events". PLOS ONE. 7 (3). e34288. Bibcode:2012PLoSO...734288H. doi:10.1371/journal.pone.0034288. PMC 3314501. PMID 22470552.
Karafet, Tatiana M.; Mendez, Fernando L.; Sudoyo, Herawati; Lansing, J. Stephen; Hammer, Michael F. (2014). "Improved phylogenetic resolution and rapid diversification of Y-chromosome haplogroup K-M526 in Southeast Asia". Nature. 23 (3): 369–373. doi:10.1038/ejhg.2014.106. PMC 4326703. PMID 24896152.
Kasperaviciūte, D.; Kucinskas, V.; Stoneking, M. (2005). "Y Chromosome and Mitochondrial DNA Variation in Lithuanians". Annals of Human Genetics. 68 (5): 438–452. doi:10.1046/j.1529-8817.2003.00119.x. PMID 15469421. S2CID 26562505.
Kayser M, Lao O, Anslinger K, Augustin C, Bargel G, Edelmann J, Elias S, Heinrich M, et al. (2005). "Significant genetic differentiation between Poland and Germany follows present-day political borders, as revealed by Y-chromosome analysis" (PDF). Human Genetics. 117 (5): 428–443. doi:10.1007/s00439-005-1333-9. PMID 15959808. S2CID 11066186. Archived from the original (PDF) on March 4, 2009.
Keyser, Christine; Bouakaze, Caroline; Crubézy, Eric; Nikolaev, Valery G.; Montagnon, Daniel; Reis, Tatiana; Ludes, Bertrand (2009). "Ancient DNA provides new insights into the history of south Siberian Kurgan people". Human Genetics. 126 (3): 395–410. doi:10.1007/s00439-009-0683-0. PMID 19449030. S2CID 21347353.
Kim, Kijeong; Brenner, Charles H.; Mair, Victor H.; Lee, Kwang-Ho; Kim, Jae-Hyun; Gelegdorj, Eregzen; Batbold, Natsag; Song, Yi-Chung; et al. (2010). "A western Eurasian male is found in 2000-year-old elite Xiongnu cemetery in Northeast Mongolia". American Journal of Physical Anthropology. 142 (3): 429–440. doi:10.1002/ajpa.21242. PMID 20091844.
Kivisild, T; Rootsi, S; Metspalu, M; Mastana, S; Kaldma, K; Parik, J; Metspalu, E; Adojaan, M; et al. (2003). "The Genetic Heritage of the Earliest Settlers Persists Both in Indian Tribal and Caste Populations". AJHG. 72 (2): 313–32. doi:10.1086/346068. PMC 379225. PMID 12536373.
Lazaridis, Iosif; et al. (2016). "Genomic insights into the origin of farming in the ancient Near East". Nature. 536 (7617): 419–424. Bibcode:2016Natur.536..419L. doi:10.1038/nature19310. PMC 5003663. PMID 27459054.
Lell JT, Sukernik RI, Starikovskaya YB, Su B, Jin L, Schurr TG, Underhill PA, Wallace DC (2002). "The Dual Origin and Siberian Affinities of Native American Y Chromosomes" (PDF). American Journal of Human Genetics. 70 (1): 192–206. doi:10.1086/338457. PMC 384887. PMID 11731934. Archived from the original (PDF) on April 22, 2003.
Liu Shu-hu; Nizam Yilihamu; Rabiyamu Bake; Abdukeram Bupatima; Dolkun Matyusup (2018). "A study of genetic diversity of three isolated populations in Xinjiang using Y-SNP". Acta Anthropologica Sinica. 37 (1): 146–156.
Carlos Quiles (September 10, 2018). "A study of genetic diversity of three isolated populations in Xinjiang using Y-SNP". Indo-European. Archived from the original on September 20, 2020. Retrieved July 24, 2020.
Malmström, Helena; Günther, Torsten; Svensson, Emma M.; Juras, Anna; Fraser, Magdalena; Munters, Arielle R.; Pospieszny, Łukasz; Tõrv, Mari; et al. (October 9, 2019). "The genomic ancestry of the Scandinavian Battle Axe Culture people and their relation to the broader Corded Ware horizon". Proceedings of the Royal Society B. 286 (1912). doi:10.1098/rspb.2019.1528. PMC 6790770. PMID 31594508.
Malyarchuk, Boris; Derenko, Miroslava; Wozniak, Marcin; Grzybowski, Tomasz (2013). "Y-chromosome variation in Tajiks and Iranians". Annals of Human Biology. 40 (1): 48–54. doi:10.3109/03014460.2012.747628. PMID 23198991. S2CID 2752490.
Mascarenhas, Desmond D.; Raina, Anupuma; Aston, Christopher E.; Sanghera, Dharambir K. (2015). "Genetic and Cultural Reconstruction of the Migration of an Ancient Lineage". BioMed Research International. 2015: 651415. doi:10.1155/2015/651415. PMC 4605215. PMID 26491681.
Mathieson, Iain; Lazaridis, Iosif; Rohland, Nadin; Mallick, Swapan; Patterson, Nick; Alpaslan Roodenberg, Songul; Harney, Eadaoin; Stewardson, Kristin; et al. (2015). "Eight thousand years of natural selection in Europe". bioRxiv10.1101/016477.
Mirabal, Sheyla; Regueiro, M; Cadenas, AM; Cavalli-Sforza, LL; Underhill, PA; Verbenko, DA; Limborska, SA; Herrera, RJ; et al. (2009). "Y-Chromosome distribution within the geo-linguistic landscape of northwestern Russia". European Journal of Human Genetics. 17 (10): 1260–1273. doi:10.1038/ejhg.2009.6. PMC 2986641. PMID 19259129.
Mohammad T, Xue Y, Evison M, Tyler-Smith C (2009). "Genetic structure of nomadic Bedouin from Kuwait". Heredity. 103 (5): 425–433. doi:10.1038/hdy.2009.72. PMC 2869035. PMID 19639002.
Narasimhan, Vagheesh M.; Anthony, David; Mallory, James; Reich, David (2018). "The Genomic Formation of South and Central Asia". bioRxiv10.1101/292581.
Narasimhan, Vagheesh M.; Patterson, N.J.; Moorjani, Priya; Rohland, Nadin; et al. (2019), "The Formation of Human Populations in South and Central Asia", Science, 365 (6457): eaat7487, doi:10.1126/science.aat7487, PMC 6822619, PMID 31488661
Nasidze I, Ling EY, Quinque D, Dupanloup I, Cordaux R, Rychkov S, Naumova O, Zhukova O, et al. (2004). "Mitochondrial DNA and Y-Chromosome Variation in the Caucasus" (PDF). Annals of Human Genetics. 68 (Pt 3): 205–221. doi:10.1046/j.1529-8817.2004.00092.x. PMID 15180701. S2CID 27204150. Archived from the original (PDF) on October 30, 2004.
Nasidze I, Quinque D, Ozturk M, Bendukidze N, Stoneking M (2005). "MtDNA and Y-chromosome Variation in Kurdish Groups" (PDF). Annals of Human Genetics. 69 (Pt 4): 401–412. doi:10.1046/j.1529-8817.2005.00174.x. PMID 15996169. S2CID 23771698. Archived from the original (PDF) on August 23, 2009.
Pamjav, Horolma; Fehér, Tibor; Németh, Endre; Pádár, Zsolt (2012), "Brief communication: new Y-chromosome binary markers improve phylogenetic resolution within haplogroup R1a1", American Journal of Physical Anthropology, 149 (4): 611–615, doi:10.1002/ajpa.22167, PMID 23115110, S2CID 4820868
Passarino G, Cavalleri GL, Lin AA, Cavalli-Sforza LL, Børresen-Dale AL, Underhill (2002). "Different genetic components in the Norwegian population revealed by the analysis of mtDNA and Y chromosome polymorphisms". European Journal of Human Genetics. 10 (9): 521–529. doi:10.1038/sj.ejhg.5200834. PMID 12173029.
Pathak, Ajai K.; Kadian, Anurag; Kushniarevich, Alena; Montinaro, Francesco; Mondal, Mayukh; Ongaro, Linda; Singh, Manvendra; Kumar, Pramod; et al. (December 6, 2018). "The Genetic Ancestry of Modern Indus Valley Populations from Northwest India". The American Journal of Human Genetics. 103 (6): 918–929. doi:10.1016/j.ajhg.2018.10.022. PMC 6288199. PMID 30526867.
Pawlowski, R; Dettlaff-Kakol, A; MacIejewska, A; Paszkowska, R; Reichert, M; Jezierski, G (2002). "Population genetics of 9 Y-chromosome STR loci w Northern Poland". Arch. Med. Sadowej Kryminol. 52 (4): 261–277. PMID 14669672.
Pericić M, Lauc LB, Klarić IM, Rootsi S, Janićijević B, Rudan I, Terzić R, Colak I, et al. (2005). "High-resolution phylogenetic analysis of southeastern Europe traces major episodes of paternal gene flow among Slavic populations". Mol. Biol. Evol. 22 (10): 1964–75. doi:10.1093/molbev/msi185. PMID 15944443.
Poznik GD, et al. (2016). "Punctuated bursts in human male demography inferred from 1,244 worldwide Y-chromosome sequences". Nature Genetics. 48 (6): 593–599. doi:10.1038/ng.3559. hdl:11858/00-001M-0000-002A-F024-C. PMC 4884158. PMID 27111036.
Ricaut F, Keyser-Tracqui C, Bourgeois I, Crubézy E, Ludes B (2004). "Genetic Analysis of a Scytho-Siberian Skeleton and Its Implications for Ancient Central Asian Migrations". Human Biology. 76 (1): 109–25. doi:10.1353/hub.2004.0025. PMID 15222683. S2CID 35948291.
Rosser ZH, Zerjal T, Hurles ME, Adojaan M, Alavantic D, Amorim A, Amos W, Armenteros M, et al. (2000). "Y-Chromosomal Diversity in Europe Is Clinal and Influenced Primarily by Geography, Rather than by Language". American Journal of Human Genetics. 67 (6): 1526–1543. doi:10.1086/316890. PMC 1287948. PMID 11078479.
Saag, Lehti; Varul, Liivi; Scheib, Christiana Lyn; Stenderup, Jesper; Allentoft, Morten E.; Saag, Lauri; Pagani, Luca; Reidla, Maere; et al. (July 24, 2017). "Extensive Farming in Estonia Started through a Sex-Biased Migration from the Steppe". Current Biology. 27 (14). Cell Press: 2185–2193. Bibcode:2017CBio...27E2185S. doi:10.1016/j.cub.2017.06.022. PMID 28712569.
Saag, Lehti; Vasilyev, Sergey V.; Varul, Liivi; Kosorukova, Natalia V.; Gerasimov, Dmitri V.; Oshibkina, Svetlana V.; Griffith, Samuel J.; Solnik, Anu; et al. (July 3, 2020). "Genetic ancestry changes in Stone to Bronze Age transition in the East European plain". bioRxiv10.1101/2020.07.02.184507.
Sahoo, S; Singh, A; Himabindu, G; Banerjee, J; Sitalaximi, T; Gaikwad, S; Trivedi, R; Endicott, P; et al. (2006). "A prehistory of Indian Y chromosomes: Evaluating demic diffusion scenarios". Proceedings of the National Academy of Sciences. 103 (4): 843–848. Bibcode:2006PNAS..103..843S. doi:10.1073/pnas.0507714103. PMC 1347984. PMID 16415161.
Scozzari R, Cruciani F, Pangrazio A, Santolamazza P, Vona G, Moral P, Latini V, Varesi L, et al. (2001). "Human Y-Chromosome Variation in the Western Mediterranean Area: Implications for the Peopling of the Region" (PDF). Human Immunology. 62 (9): 871–84. CiteSeerX10.1.1.408.4857. doi:10.1016/S0198-8859(01)00286-5. PMID 11543889. Archived from the original (PDF) on December 17, 2012. Retrieved March 15, 2009.
Semenov, Alexander S.; Bulat, Vladimir V. (2016), "Ancient Paleo-DNA of Pre-Copper Age North-Eastern Europe: Establishing the Migration Traces of R1a1 Y-DNA Haplogroup", European Journal of Molecular Biotechnology, 11 (1): 40–54, doi:10.13187/ejmb.2016.11.40, S2CID 172131289
Semino, O; Passarino, G; Oefner, PJ; Lin, AA; Arbuzova, S; Beckman, LE; De Benedictis, G; Francalacci, P; et al. (2000). "The Genetic Legacy of Paleolithic Homo sapiens sapiens in Extant Europeans: A Y Chromosome Perspective" (PDF). Science. 290 (5494): 1155–1159. Bibcode:2000Sci...290.1155S. doi:10.1126/science.290.5494.1155. PMID 11073453. Archived from the original (PDF) on November 25, 2003.
Sengupta, S; Zhivotovsky, LA; King, R; Mehdi, SQ; Edmonds, CA; Chow, CE; Lin, AA; Mitra, M; et al. (2006). "Polarity and Temporality of High-Resolution Y-Chromosome Distributions in India Identify Both Indigenous and Exogenous Expansions and Reveal Minor Genetic Influence of Central Asian Pastoralists". American Journal of Human Genetics. 78 (2): 202–21. doi:10.1086/499411. PMC 1380230. PMID 16400607.
Sharma, S; Rai, E; Sharma, P; Jena, M; Singh, S; Darvishi, K; Bhat, AK; Bhanwer, AJ; et al. (2009). "The Indian origin of paternal haplogroup R1a1(*)substantiates the autochthonous origin of Brahmins and the caste system". Journal of Human Genetics. 54 (1): 47–55. doi:10.1038/jhg.2008.2. PMID 19158816.
Schilz, Felix (2006). Molekulargenetische Verwandtschaftsanalysen am prähistorischen Skelettkollektiv der Lichtensteinhöhle [Molecular genetic kinship analysis on the prehistoric skeleton collective of the Lichtenstein Cave] (PDF) (Dissertation) (in German). Göttingen: Mathematisch-Naturwissenschaftlichen Fakultäten der Georg-August-Universität. Archived (PDF) from the original on June 8, 2011. Retrieved November 9, 2009.
Silva, Marina; et al. (2017). "A genetic chronology for the Indian Subcontinent points to heavily sex-biased dispersals". BMC Evolutionary Biology. 17 (1): 88. Bibcode:2017BMCEE..17...88S. doi:10.1186/s12862-017-0936-9. PMC 5364613. PMID 28335724.
Tambets K, Rootsi S, Kivisild T, Help H, Serk P, Loogväli EL, Tolk HV, Reidla M, et al. (2004). "The Western and Eastern Roots of the Saami—the Story of Genetic 'Outliers' Told by Mitochondrial DNA and Y Chromosomes". American Journal of Human Genetics. 74 (4): 661–682. doi:10.1086/383203. PMC 1181943. PMID 15024688.
Thangaraj, Kumarasamy; Naidu, B. Prathap; Crivellaro, Federica; Tamang, Rakesh; Upadhyay, Shashank; Sharma, Varun Kumar; Reddy, Alla G.; Walimbe, S. R.; et al. (2010). Cordaux, Richard (ed.). "The Influence of Natural Barriers in Shaping the Genetic Structure of Maharashtra Populations". PLOS ONE. 5 (12): e15283. Bibcode:2010PLoSO...515283T. doi:10.1371/journal.pone.0015283. PMC 3004917. PMID 21187967.
Underhill, PA; Myres, NM; Rootsi, S; Metspalu, M; Zhivotovsky, LA; King, RJ; Lin, AA; Chow, CE; et al. (November 4, 2009). "Separating the post-Glacial coancestry of European and Asian Y chromosomes within haplogroup R1a". European Journal of Human Genetics. 18 (4) (published April 2010): 479–84. doi:10.1038/ejhg.2009.194. PMC 2987245. PMID 19888303.
Underhill, Peter A.; et al. (March 26, 2014). "The phylogenetic and geographic structure of Y-chromosome haplogroup R1a". European Journal of Human Genetics. 23 (1) (published January 2015): 124–131. doi:10.1038/ejhg.2014.50. PMC 4266736. PMID 24667786. "PDF" (PDF). Archived from the original (PDF) on August 16, 2016. Retrieved June 12, 2016.
Wang, Wei; Wise, Cheryl; Baric, Tom; Black, Michael L.; Bittles, Alan H. (2003). "The origins and genetic structure of three co-resident Chinese Muslim populations: The Salar, Bo'an and Dongxiang". Human Genetics. 113 (3): 244–52. doi:10.1007/s00439-003-0948-y. PMID 12759817. S2CID 11138499.
Wells, R.S. (2001), "The Eurasian Heartland: A continental perspective on Y-chromosome diversity", Proceedings of the National Academy of Sciences of the USA, 98 (18): 10244–10249, Bibcode:2001PNAS...9810244W, doi:10.1073/pnas.171305098, PMC 56946, PMID 11526236
Yan, Shi; Wang, Chuan-Chao; Zheng, Hong-Xiang; Wang, Wei; Qin, Zhen-Dong; Wei, Lan-Hai; Wang, Yi; Pan, Xue-Dong; et al. (August 29, 2014). "Y Chromosomes of 40% Chinese Descend from Three Neolithic Super-Grandfathers". PLOS ONE. 9 (8). e105691. arXiv:1310.3897. Bibcode:2014PLoSO...9j5691Y. doi:10.1371/journal.pone.0105691. PMC 4149484. PMID 25170956.
Y Chromosome Consortium "YCC" (2002). "A Nomenclature System for the Tree of Human Y-Chromosomal Binary Haplogroups". Genome Research. 12 (2): 339–348. doi:10.1101/gr.217602. PMC 155271. PMID 11827954.
Zerjal, Tatiana; Wells, R. Spencer; Yuldasheva, Nadira; Ruzibakiev, Ruslan; Tyler-Smith, Chris (2002). "A Genetic Landscape Reshaped by Recent Events: Y-Chromosomal Insights into Central Asia". The American Journal of Human Genetics. 71 (3): 466–82. doi:10.1086/342096. PMC 419996. PMID 12145751.
Zhong H, Shi H, Qi XB, Duan Y, Tan PP, Jin L, SU B, Ma RZ (January 2011). "Extended Y chromosome investigation suggests postglacial migrations of modern humans into East Asia via the northern route". Molecular Biology and Evolution. 28 (1): 717–27. doi:10.1093/molbev/msq247. PMID 20837606.
Zhou, Ruixia; An, Lizhe; Wang, Xunling; Shao, Wei; Lin, Gonghua; Yu, Weiping; Yi, Lin; Xu, Shijian; et al. (2007). "Testing the hypothesis of an ancient Roman soldier origin of the Liqian people in northwest China: a Y-chromosome perspective". Journal of Human Genetics. 52 (7): 584–91. doi:10.1007/s10038-007-0155-0. PMID 17579807.
Further reading
Gimbutas (1970). Indo-European and Indo-Europeans. Philadelphia, PA: Univ. of Pennsylvania Press. pp. 155–195.
Patrizia Malaspina; Andrey I. Kozlov; Fulvio Cruciani; Piero Santolamazza; Nejat Akar; Dimiter Kovatchev; Marina G. Kerimova; Juri Parik; Richard Villems; Rosana Scozzari; Andrea Novelletto (2003). "Analysis of Y-chromosome variation in modern populations at the European-Asian border" (PDF). In K. Boyle; C. Renfrew; M. Levine (eds.). Ancient interactions: east and west in Eurasia. McDonald Institute Monographs. Cambridge: Cambridge University Press. pp. 309–313. Archived from the original (PDF) on June 24, 2023. Retrieved September 10, 2009.
Parpola, Asko (2015). The Roots of Hinduism. The early Aryans and the Indus Civilisation. Oxford University Press.
Sharma, S. (2007). "1344/T: The Autochthonous Origin and a Tribal Link of Indian Brahmins: Evaluation Through Molecular Genetic Markers". The American Society of Human Genetics 57th Annual Meeting October 23–27, 2007; San Diego, California (PDF) (Abstract). p. 273. Archived from the original (PDF) on June 26, 2008.
Trautmann, Thomas (2005). The Aryan Debate. Oxford University Press.
Varzari, Alexander (2006). Population History of the Dniester-Carpathians: Evidence from Alu Insertion and Y-Chromosome Polymorphisms (PDF) (Dissertation). München: Ludwig-Maximilians-Universität. Archived (PDF) from the original on October 28, 2008. Retrieved October 30, 2008.

_and_R1a1a_oldest_expansion_and_highest_frequency_(2014).jpg/1280px-R1a_origins_(Underhill_2010)_and_R1a1a_oldest_expansion_and_highest_frequency_(2014).jpg)