La nitrificación es la oxidación biológica del amoníaco a nitrato a través del intermediario nitrito . La nitrificación es un paso importante en el ciclo del nitrógeno en el suelo . El proceso de nitrificación completa puede ocurrir a través de organismos separados [1] o completamente dentro de un organismo, como en las bacterias comammox . La transformación del amoníaco a nitrito suele ser el paso limitante de la velocidad de la nitrificación. La nitrificación es un proceso aeróbico realizado por pequeños grupos de bacterias autótrofas y arqueas .
El proceso de nitrificación comienza con la primera etapa de oxidación del amoniaco, donde el amoniaco (NH 3 ) o el amonio (NH 4 + ) se convierten en nitrito (NO 2 - ). Esta primera etapa a veces se conoce como nitritación. La llevan a cabo dos grupos de organismos, las bacterias oxidantes del amoniaco ( AOB ) y las arqueas oxidantes del amoniaco ( AOA [2] ).
Las bacterias oxidantes de amoníaco (AOB) son típicamente bacterias gramnegativas y pertenecen a Betaproteobacteria y Gammaproteobacteria [3], incluidos los géneros comúnmente estudiados, como Nitrosomonas y Nitrococcus . Son conocidas por su capacidad de utilizar el amoníaco como fuente de energía y prevalecen en una amplia gama de entornos, como suelos, sistemas acuáticos y plantas de tratamiento de aguas residuales.
Las AOB poseen enzimas llamadas amoniaco monooxigenasas (AMO), que son responsables de catalizar la conversión de amoniaco en hidroxilamina (NH 2 OH), un intermediario crucial en el proceso de nitrificación. [4] Esta actividad enzimática es sensible a factores ambientales, como el pH, la temperatura y la disponibilidad de oxígeno.
Las AOB desempeñan un papel vital en la nitrificación del suelo, lo que las convierte en actores clave en el ciclo de nutrientes . Contribuyen a la transformación del amoníaco derivado de la descomposición de materia orgánica o fertilizantes en nitrito, que posteriormente sirve como sustrato para las bacterias oxidantes de nitrito (NOB).
Antes del descubrimiento de arqueas capaces de oxidar amoníaco, las bacterias oxidantes de amoníaco (AOB) se consideraban los únicos organismos capaces de oxidar amoníaco. Desde su descubrimiento en 2005, [5] se han cultivado dos aislados de AOA: Nitrosopumilus maritimus [6] y Nitrososphaera viennensis . [7] Al comparar AOB y AOA, AOA domina tanto en suelos como en ambientes marinos, [2] [8] [6] [9] [10] [11] lo que sugiere que Nitrososphaerota (anteriormente Thaumarchaeota ) puede ser un mayor contribuyente a la oxidación de amoníaco en estos ambientes. [2]
El crenarchaeol , que generalmente se cree que es producido exclusivamente por AOA (específicamente Nitrososphaerota), se ha propuesto como un biomarcador para AOA y oxidación de amoníaco. Se ha descubierto que la abundancia de crenarchaeol sigue las floraciones estacionales de AOA, lo que sugiere que puede ser apropiado usar las abundancias de crenarchaeol como un indicador de las poblaciones de AOA [12] y, por lo tanto, de la oxidación de amoníaco en general. Sin embargo, el descubrimiento de Nitrososphaerota que no son oxidantes obligados de amoníaco [13] complica esta conclusión, [14] al igual que un estudio que sugiere que el crenarchaeol puede ser producido por Euryarchaeota del Grupo II Marino. [15]
El segundo paso de la nitrificación es la oxidación del nitrito en nitrato. Este proceso a veces se conoce como nitratación. La oxidación del nitrito es realizada por bacterias oxidantes de nitrito ( BNO ) de los taxones Nitrospirota , [16] Nitrospinota , [17] Pseudomonadota [18] y Chloroflexota . [19] Las BNO suelen estar presentes en el suelo, en manantiales geotermales, en agua dulce y en ecosistemas marinos.
La oxidación del amoniaco a nitrato en un solo paso dentro de un organismo fue predicha en 2006 [20] y descubierta en 2015 en la especie Nitrospira inopinata . En 2017 se obtuvo un cultivo puro del organismo [21] , lo que representa una revolución en nuestra comprensión del proceso de nitrificación.
La idea de que la oxidación del amoniaco a nitrato es de hecho un proceso biológico fue dada por primera vez por Louis Pasteur en 1862. [22] Más tarde, en 1875, Alexander Müller, mientras realizaba una evaluación de calidad del agua de los pozos en Berlín , notó que el amonio era estable en soluciones esterilizadas pero nitrificado en aguas naturales. A. Müller propuso que la nitrificación la realizan microorganismos. [23] En 1877, Jean-Jacques Schloesing y Achille Müntz , dos químicos agrícolas franceses que trabajaban en París , demostraron que la nitrificación es de hecho un proceso mediado por microbios mediante experimentos con aguas residuales líquidas y matriz de suelo artificial (arena esterilizada con tiza en polvo). [24] Sus hallazgos fueron confirmados pronto (en 1878) por Robert Warington , quien estaba investigando la capacidad de nitrificación del suelo de jardín en la estación experimental de Rothamsted en Harpenden en Inglaterra. [25] R. Warington también hizo la primera observación de que la nitrificación es un proceso de dos pasos en 1879 [26] que fue confirmada por John Munro en 1886. [27] Aunque en ese momento, se creía que la nitrificación de dos pasos se separa en fases de vida distintas o rasgos característicos de un solo microorganismo.
El primer nitrificador puro (oxidante de amoniaco) fue probablemente aislado en 1890 por Percy Frankland y Grace Frankland , dos científicos ingleses de Escocia. [28] Antes de eso, Warington , [25] Sergei Winogradsky [29] y los Frankland solo pudieron enriquecer cultivos de nitrificantes. Frankland y Frankland tuvieron éxito con un sistema de diluciones en serie con un inóculo muy bajo y tiempos de cultivo largos que contaban en años. Sergei Winogradsky afirmó haber aislado un cultivo puro en el mismo año (1890), [29] pero su cultivo seguía siendo un co-cultivo de bacterias oxidantes de amoniaco y nitrito. [30] S. Winogradsky tuvo éxito solo un año después, en 1891. [31]
De hecho, durante las diluciones seriadas, los oxidantes de amoníaco y los oxidantes de nitrito se separaron sin saberlo, lo que dio como resultado un cultivo puro con capacidad de oxidación de amoníaco únicamente. Por lo tanto, Frankland y Frankland observaron que estos cultivos puros pierden la capacidad de realizar ambos pasos. La pérdida de la capacidad de oxidación de nitrito ya fue observada por R. Warington . [26] El cultivo de oxidante de nitrito puro ocurrió más tarde durante el siglo XX, sin embargo, no es posible estar seguro de qué cultivos estaban libres de contaminantes ya que todas las cepas teóricamente puras comparten el mismo rasgo (consumo de nitrito, producción de nitrato). [30]
Ambos pasos producen energía que se acopla a la síntesis de ATP. Los organismos nitrificantes son quimioautótrofos y utilizan dióxido de carbono como fuente de carbono para el crecimiento. Algunos AOB poseen la enzima ureasa , que cataliza la conversión de la molécula de urea en dos moléculas de amoníaco y una molécula de dióxido de carbono. Se ha demostrado que Nitrosomonas europaea , así como las poblaciones de AOB que viven en el suelo, asimilan el dióxido de carbono liberado por la reacción para producir biomasa a través del ciclo de Calvin y obtienen energía oxidando amoníaco (el otro producto de la ureasa) a nitrito. Esta característica puede explicar el mayor crecimiento de AOB en presencia de urea en entornos ácidos. [32]
En la mayoría de los ambientes, existen organismos que completarán ambos pasos del proceso, obteniendo nitrato como producto final. Sin embargo, es posible diseñar sistemas en los que se forme nitrito (el proceso Sharon ).
La nitrificación es importante en los sistemas agrícolas, donde el fertilizante se aplica a menudo en forma de amoníaco. La conversión de este amoníaco en nitrato aumenta la lixiviación de nitrógeno porque el nitrato es más soluble en agua que el amoníaco.
La nitrificación también desempeña un papel importante en la eliminación de nitrógeno de las aguas residuales municipales . La eliminación convencional es la nitrificación, seguida de la desnitrificación . El coste de este proceso reside principalmente en la aireación (llevar oxígeno al reactor) y la adición de una fuente externa de carbono (por ejemplo, metanol ) para la desnitrificación.
La nitrificación también puede ocurrir en el agua potable. En los sistemas de distribución donde se utilizan cloraminas como desinfectante secundario, la presencia de amoníaco libre puede actuar como sustrato para los microorganismos oxidantes del amoníaco. Las reacciones asociadas pueden conducir al agotamiento del desinfectante residual en el sistema. [33] Se ha demostrado que la adición de iones clorito al agua tratada con cloramina controla la nitrificación. [34] [35]
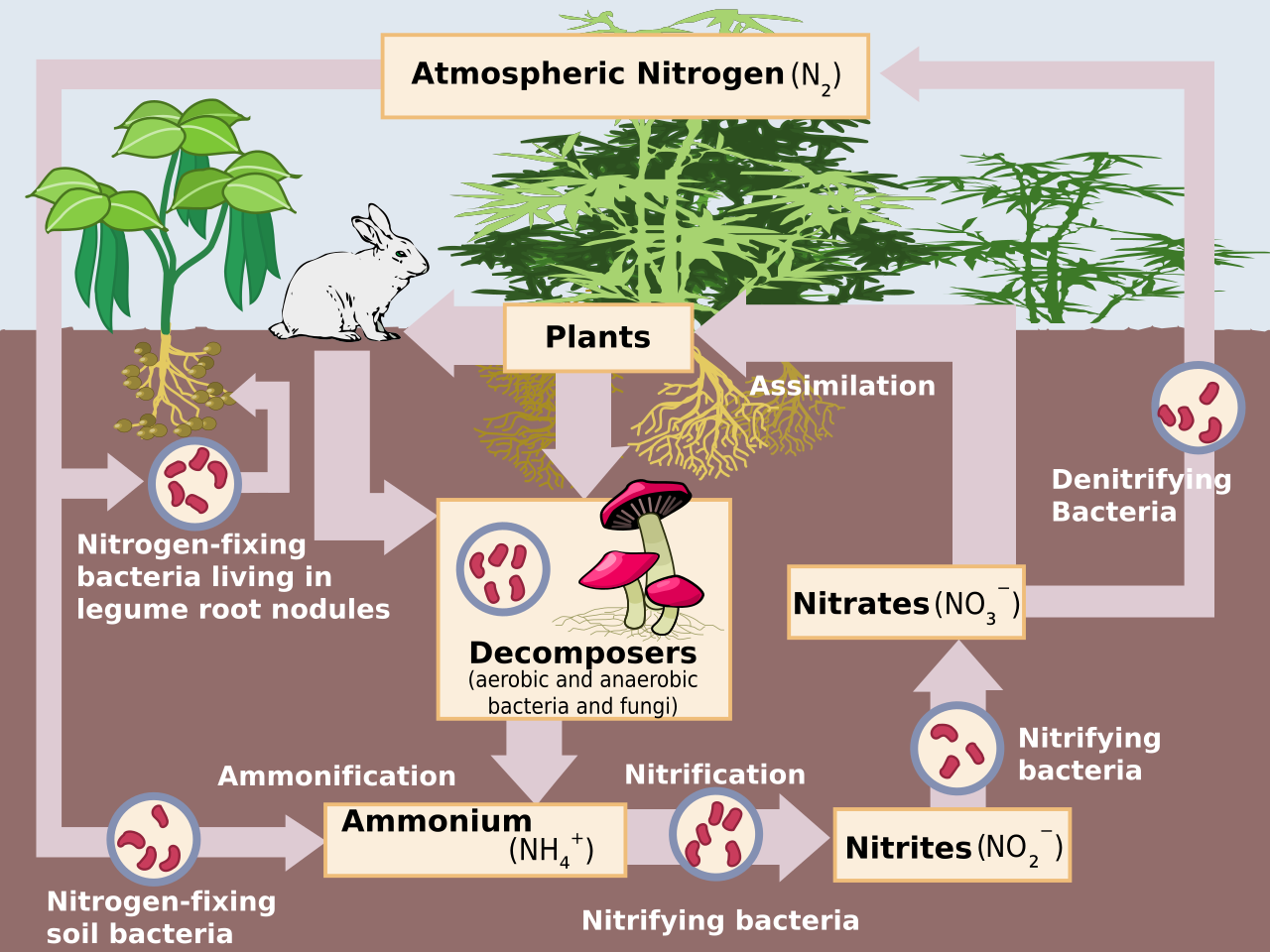
Junto con la amonificación , la nitrificación forma un proceso de mineralización que se refiere a la descomposición completa de la materia orgánica, con la liberación de compuestos nitrogenados disponibles. Esto repone el ciclo del nitrógeno .
En el ambiente marino , el nitrógeno es a menudo el nutriente limitante , por lo que el ciclo del nitrógeno en el océano es de particular interés. [36] [37] El paso de nitrificación del ciclo es de particular interés en el océano porque crea nitrato , la forma primaria de nitrógeno responsable de la "nueva" producción . Además, a medida que el océano se enriquece con CO 2 antropogénico , la disminución resultante del pH podría conducir a una disminución de las tasas de nitrificación. La nitrificación podría convertirse potencialmente en un "cuello de botella" en el ciclo del nitrógeno. [38]
La nitrificación, como se indicó anteriormente, es formalmente un proceso de dos pasos; en el primer paso, el amoníaco se oxida a nitrito , y en el segundo paso, el nitrito se oxida a nitrato. Diversos microbios son responsables de cada paso en el entorno marino. Se conocen varios grupos de bacterias oxidantes de amoníaco (AOB) en el entorno marino, incluidas Nitrosomonas , Nitrospira y Nitrosococcus . Todas contienen el gen funcional amoniaco monooxigenasa ( AMO ) que, como su nombre lo indica, es responsable de la oxidación del amoníaco. [2] [37] Estudios metagenómicos posteriores y enfoques de cultivo han revelado que algunos Thermoproteota (anteriormente Crenarchaeota) poseen AMO. Thermoproteota son abundantes en el océano y algunas especies tienen una afinidad 200 veces mayor por el amoníaco que las AOB, lo que contrasta con la creencia anterior de que las AOB son las principales responsables de la nitrificación en el océano. [39] [36] Además, aunque clásicamente se piensa que la nitrificación está separada verticalmente de la producción primaria porque la oxidación del nitrato por las bacterias es inhibida por la luz, la nitrificación por AOA no parece ser inhibida por la luz, lo que significa que la nitrificación está ocurriendo en toda la columna de agua , desafiando las definiciones clásicas de producción "nueva" y "reciclada" . [36]
En el segundo paso, el nitrito se oxida a nitrato. En los océanos, este paso no se comprende tan bien como el primero, pero se sabe que las bacterias Nitrospina [17] [40] y Nitrobacter llevan a cabo este paso en el océano. [36]
La nitrificación es un proceso de oxidación de compuestos de nitrógeno (efectivamente, pérdida de electrones del átomo de nitrógeno a los átomos de oxígeno ), y es catalizada paso a paso por una serie de enzimas.
O
En Nitrosomonas europaea , el primer paso de oxidación (amoniaco a hidroxilamina ) lo lleva a cabo la enzima amoniaco monooxigenasa (AMO).
El segundo paso (transformación de la hidroxilamina en nitrito) es catalizado por dos enzimas. La hidroxilamina oxidorreductasa (HAO) convierte la hidroxilamina en óxido nítrico. [41]
Otra enzima actualmente desconocida convierte el óxido nítrico en nitrito.
El tercer paso (de nitrito a nitrato) se completa en un organismo distinto.
Debido a su naturaleza microbiana inherente, la nitrificación en los suelos es muy susceptible a las condiciones del suelo. En general, la nitrificación del suelo se producirá a un ritmo óptimo si las condiciones de las comunidades microbianas fomentan el crecimiento y la actividad microbiana saludables. Las condiciones del suelo que tienen un efecto sobre las tasas de nitrificación incluyen:
Los inhibidores de la nitrificación son compuestos químicos que retardan la nitrificación del amoníaco , fertilizantes que contienen amonio o urea , que se aplican al suelo como fertilizantes. Estos inhibidores pueden ayudar a reducir las pérdidas de nitrógeno en el suelo que de otro modo serían utilizadas por los cultivos. Los inhibidores de la nitrificación se utilizan ampliamente y se añaden a aproximadamente el 50% del amoníaco anhidro aplicado en otoño en estados de los EE. UU., como Illinois. [42] Por lo general, son eficaces para aumentar la recuperación de fertilizantes nitrogenados en cultivos en hileras, pero el nivel de eficacia depende de las condiciones externas y es más probable que sus beneficios se vean con tasas de nitrógeno inferiores a las óptimas. [43]
Las preocupaciones ambientales de la nitrificación también contribuyen al interés en el uso de inhibidores de la nitrificación: el producto principal, el nitrato , se filtra en las aguas subterráneas, produciendo toxicidad tanto en los seres humanos [44] como en algunas especies de vida silvestre y contribuyendo a la eutrofización del agua estancada. Algunos inhibidores de la nitrificación también inhiben la producción de metano , un gas de efecto invernadero.
La inhibición del proceso de nitrificación se facilita principalmente por la selección e inhibición/destrucción de las bacterias que oxidan los compuestos de amoniaco. Una multitud de compuestos inhiben la nitrificación, que se pueden dividir en las siguientes áreas: el sitio activo de la amonia monooxigenasa (AMO), inhibidores mecanísticos y el proceso de compuestos N-heterocíclicos . El proceso para el último de los tres aún no se entiende ampliamente, pero es prominente. La presencia de AMO se ha confirmado en muchos sustratos que son inhibidores de nitrógeno como diciandiamida , tiosulfato de amonio y nitrapirina .
La conversión de amoníaco en hidroxilamina es el primer paso en la nitrificación, donde AH2 representa una gama de potenciales donadores de electrones.
Esta reacción es catalizada por la AMO. Los inhibidores de esta reacción se unen al sitio activo de la AMO y previenen o retrasan el proceso. El proceso de oxidación del amoniaco por la AMO se considera importante debido a que otros procesos requieren la cooxidación de NH3 para obtener equivalentes reductores. Esto suele ser proporcionado por el compuesto hidroxilamina oxidorreductasa (HAO), que cataliza la reacción:
El mecanismo de inhibición se complica por este requisito. El análisis cinético de la inhibición de la oxidación del NH 3 ha demostrado que los sustratos de AMO han mostrado una cinética que va desde competitiva a no competitiva . La unión y la oxidación pueden ocurrir en dos sitios en AMO: en sustratos competitivos, la unión y la oxidación ocurren en el sitio del NH 3 , mientras que en sustratos no competitivos ocurre en otro sitio.
Los inhibidores basados en mecanismos pueden definirse como compuestos que interrumpen la reacción normal catalizada por una enzima. Este método se produce por la inactivación de la enzima a través de la modificación covalente del producto, que en última instancia inhibe la nitrificación. A través del proceso, el AMO se desactiva y una o más proteínas se unen covalentemente al producto final. Se ha descubierto que esto es más prominente en una amplia gama de compuestos azufrados o acetilénicos .
Se ha descubierto que los compuestos que contienen azufre, incluido el tiosulfato de amonio (un inhibidor popular), actúan produciendo compuestos volátiles con fuertes efectos inhibidores, como el disulfuro de carbono y la tiourea .
En particular, la tiofosforil triamida ha sido una adición notable donde tiene el doble propósito de inhibir tanto la producción de ureasa como la nitrificación. [45] En un estudio de los efectos inhibidores de la oxidación por la bacteria Nitrosomonas europaea , el uso de tioéteres resultó en la oxidación de estos compuestos a sulfóxidos , donde el átomo de S es el sitio primario de oxidación por AMO. Esto está más fuertemente correlacionado con el campo de la inhibición competitiva.

Los compuestos N-heterocíclicos también son inhibidores de la nitrificación muy eficaces y a menudo se clasifican por su estructura de anillo. El modo de acción de estos compuestos no se entiende bien: mientras que la nitrapirina, un inhibidor y sustrato ampliamente utilizado de AMO, es un inhibidor débil basado en el mecanismo de dicha enzima, los efectos de dicho mecanismo no pueden correlacionarse directamente con la capacidad del compuesto para inhibir la nitrificación. Se sugiere que la nitrapirina actúa contra la enzima monooxigenasa dentro de las bacterias, previniendo el crecimiento y la oxidación de CH 4 /NH 4 . [46] Los compuestos que contienen dos o tres átomos de N en el anillo adyacentes ( piridazina , pirazol , indazol ) tienden a tener un efecto de inhibición significativamente mayor que los compuestos que contienen átomos de N no adyacentes o átomos de N en el anillo singulares ( piridina , pirrol ). [47] Esto sugiere que la presencia de átomos de N en el anillo está directamente correlacionada con el efecto de inhibición de esta clase de compuestos.
Algunos inhibidores enzimáticos de la nitrificación, como la nitrapirina, también pueden inhibir la oxidación del metano en bacterias metanotróficas . [48] La AMO muestra tasas de recambio cinético similares a la metano monooxigenasa (MMO) encontrada en los metanotrofos, lo que indica que la MMO es un catalizador similar a la AMO para el propósito de la oxidación del metano. Además, las bacterias metanotróficas comparten muchas similitudes con los oxidantes de NH 3 como Nitrosomonas . [49] El perfil inhibidor de las formas particuladas de MMO (pMMO) muestra similitud con el perfil de AMO, lo que lleva a una similitud en las propiedades entre la MMO en los metanotrofos y la AMO en los autótrofos .

Los inhibidores de la nitrificación también son de interés desde un punto de vista medioambiental debido a la producción de nitratos y óxido nitroso a partir del proceso de nitrificación. El óxido nitroso (N2O ) , aunque su concentración atmosférica es mucho menor que la del CO2 , tiene un potencial de calentamiento global unas 300 veces mayor que el dióxido de carbono y contribuye en un 6% al calentamiento planetario debido a los gases de efecto invernadero. Este compuesto también es notable por catalizar la ruptura del ozono en la estratosfera . [50] Los nitratos, un compuesto tóxico para la fauna y el ganado y un producto de la nitrificación, también son motivo de preocupación.
El suelo, que consiste en arcillas polianiónicas y silicatos , generalmente tiene una carga aniónica neta. En consecuencia, el amonio (NH 4 + ) se une firmemente al suelo, pero los iones de nitrato (NO 3 − ) no lo hacen. Debido a que el nitrato es más móvil, se filtra en los suministros de agua subterránea a través de la escorrentía agrícola . Los nitratos en el agua subterránea pueden afectar las concentraciones en el agua superficial, ya sea a través de interacciones directas entre el agua subterránea y el agua superficial (por ejemplo, al ganar tramos de arroyos, manantiales), o desde cuando se extrae para uso superficial. A modo de ejemplo, gran parte del agua potable en los Estados Unidos proviene del agua subterránea, pero la mayoría de las plantas de tratamiento de aguas residuales vierten en el agua superficial.
Entre la vida silvestre, los anfibios (renacuajos) y los huevos de peces de agua dulce son los más sensibles a los niveles elevados de nitrato, y experimentan daños en el crecimiento y el desarrollo a niveles que se encuentran comúnmente en los cuerpos de agua dulce de EE. UU. (<20 mg/l); mientras que los invertebrados de agua dulce son más tolerantes (~90+ mg/l) y los peces de agua dulce adultos pueden tolerar niveles muy altos (800 mg+/l). [51] Los niveles de nitrato también contribuyen a la eutrofización , un proceso en el que las grandes floraciones de algas reducen los niveles de oxígeno en los cuerpos de agua y provocan la muerte de criaturas que consumen oxígeno debido a la anoxia. También se cree que la nitrificación contribuye a la formación de smog fotoquímico , ozono a nivel del suelo, lluvia ácida , cambios en la diversidad de especies y otros procesos indeseables. Además, también se ha demostrado que los inhibidores de la nitrificación suprimen la oxidación del metano (CH 4 ), un potente gas de efecto invernadero , a CO 2 . Se ha demostrado que tanto la nitrapirina como el acetileno son supresores especialmente fuertes de ambos procesos, aunque no están claros los modos de acción que los distinguen.






