El reflejo vestíbulo-ocular ( VOR ) es un reflejo que actúa para estabilizar la mirada durante el movimiento de la cabeza, con el movimiento de los ojos debido a la activación del sistema vestibular , también se conoce como el reflejo cervico-ocular. El reflejo actúa para estabilizar las imágenes en las retinas del ojo durante el movimiento de la cabeza. La mirada se mantiene firme en un lugar produciendo movimientos oculares en la dirección opuesta al movimiento de la cabeza. Por ejemplo, cuando la cabeza se mueve hacia la derecha, los ojos se mueven hacia la izquierda, lo que significa que la imagen que ve una persona permanece igual aunque la cabeza se haya girado. Dado que el ligero movimiento de la cabeza está presente todo el tiempo, el VOR es necesario para estabilizar la visión: las personas con un reflejo deteriorado tienen dificultades para leer usando letra impresa, porque los ojos no se estabilizan durante pequeños temblores de la cabeza, y también porque el daño al reflejo puede causar nistagmo . [1]
El VOR no depende de lo que se ve. También puede activarse mediante estimulación con calor o frío del oído interno , donde se asienta el sistema vestibular, y funciona incluso en total oscuridad o con los ojos cerrados. Sin embargo, en presencia de luz, al movimiento también se suma el reflejo de fijación . [2] La mayoría de las características del VOR están presentes en los gatitos criados en completa oscuridad. [3]
En los animales inferiores, los órganos que coordinan el equilibrio y el movimiento no son independientes del movimiento ocular. Un pez, por ejemplo, mueve los ojos por reflejo cuando mueve la cola. Los humanos tenemos canales semicirculares , receptores de "estiramiento" de los músculos del cuello y el utrículo (órgano de la gravedad). Aunque los canales semicirculares causan la mayoría de los reflejos que responden a la aceleración, el mantenimiento del equilibrio está mediado por el estiramiento de los músculos del cuello y la atracción de la gravedad sobre el utrículo (órgano otolítico) del oído interno. [2]
El VOR tiene aspectos rotacionales y traslacionales. Cuando la cabeza rota sobre cualquier eje (horizontal, vertical o torsional), las imágenes visuales distantes se estabilizan al rotar los ojos sobre el mismo eje, pero en la dirección opuesta. [4] Cuando la cabeza se traslada, por ejemplo al caminar, el punto de fijación visual se mantiene al rotar la dirección de la mirada en la dirección opuesta, [5] en una cantidad que depende de la distancia. [6]
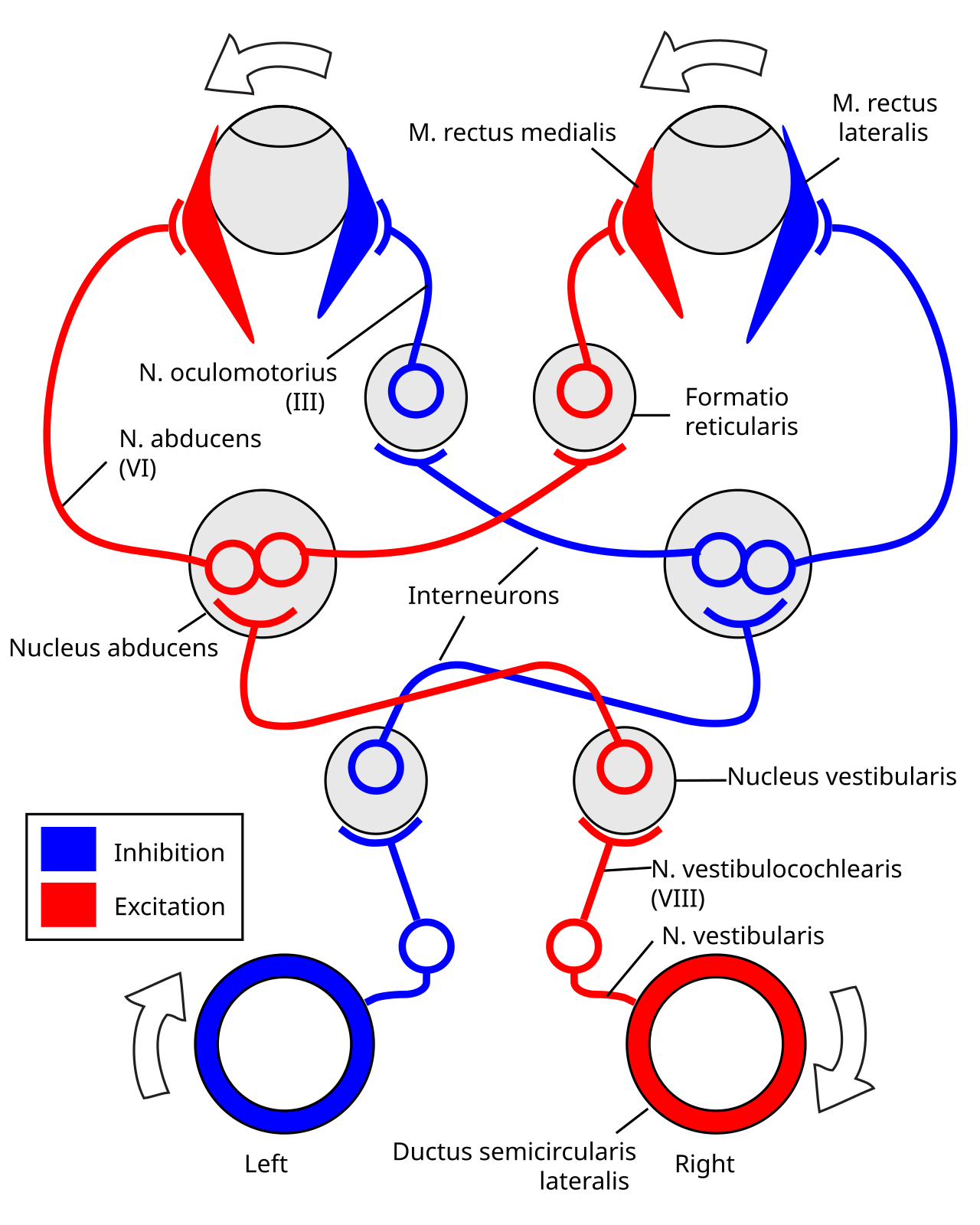
El reflejo vestíbulo-ocular es impulsado por señales que surgen del sistema vestibular del oído interno. Los canales semicirculares detectan la rotación de la cabeza y proporcionan el componente rotacional, mientras que los otolitos detectan la traslación de la cabeza y impulsan el componente traslacional. La señal para el componente rotacional horizontal viaja a través del nervio vestibular a través del ganglio vestibular y termina en los núcleos vestibulares en el tronco encefálico . Desde estos núcleos, las fibras cruzan al núcleo abducens del lado opuesto del cerebro. Aquí, las fibras hacen sinapsis con 2 vías adicionales. Una vía se proyecta directamente al músculo recto lateral del ojo a través del nervio abducens. Otro tracto nervioso se proyecta desde el núcleo abducens por el fascículo longitudinal medial al núcleo oculomotor del lado opuesto, que contiene neuronas motoras que impulsan la actividad muscular ocular, activando específicamente el músculo recto medial del ojo a través del nervio oculomotor .
Otra vía (no aparece en la imagen) se proyecta directamente desde el núcleo vestibular a través del tracto ascendente de Deiter hasta la neurona motora del músculo recto medial del mismo lado. Además, existen vías vestibulares inhibidoras hacia el núcleo abducens ipsilateral. Sin embargo, no existe una vía directa de neurona vestibular a neurona motora del músculo recto medial. [7]
Existen vías similares para los componentes verticales y torsionales del VOR.
Además de estas vías directas, que impulsan la velocidad de rotación del ojo, existe una vía indirecta que genera la señal de posición necesaria para evitar que el ojo gire hacia atrás hasta el centro cuando la cabeza deja de moverse. Esta vía es particularmente importante cuando la cabeza se mueve lentamente porque aquí las señales de posición dominan sobre las señales de velocidad. David A. Robinson descubrió que los músculos oculares requieren este impulso dual de velocidad-posición, y también propuso que debe surgir en el cerebro mediante la integración matemática de la señal de velocidad y luego enviando la señal de posición resultante a las neuronas motoras. Robinson estaba en lo cierto: el "integrador neuronal" para la posición horizontal del ojo se encontró en el núcleo prepositus hipoglossi [8] en el bulbo raquídeo, y el integrador neuronal para las posiciones verticales y torsionales del ojo se encontró en el núcleo intersticial de Cajal [9] en el mesencéfalo. Los mismos integradores neuronales también generan la posición del ojo para otros movimientos oculares conjugados, como los movimientos sacádicos y el seguimiento suave .
El integrador tiene fugas , con un tiempo de fuga característico de 20 s. Por ejemplo, cuando el sujeto está sentado quieto y concentrado en un objeto, y de repente se apaga la luz, los ojos volverían a su posición neutra en unos 40 segundos, incluso mientras el sujeto intenta mantener el foco. [10] [11]
Por ejemplo, si la cabeza se gira en el sentido de las agujas del reloj, como se ve desde arriba, los impulsos excitatorios se envían desde el canal semicircular del lado derecho a través del nervio vestibular a través del ganglio de Scarpa y terminan en los núcleos vestibulares derechos del tronco encefálico. Desde estos núcleos, las fibras excitatorias cruzan al núcleo abducens izquierdo. Allí se proyectan y estimulan el recto lateral del ojo izquierdo a través del nervio abducens. Además, a través del fascículo longitudinal medial y los núcleos oculomotores , activan los músculos rectos mediales del ojo derecho. Como resultado, ambos ojos girarán en sentido contrario a las agujas del reloj.
Además, algunas neuronas del núcleo vestibular derecho estimulan directamente las neuronas motoras del recto medial derecho e inhiben el núcleo abducens derecho.
El VOR está controlado por un integrador neuronal. La neurona de cada canal semicircular horizontal se activa a una velocidad de , donde es la velocidad angular horizontal detectada del canal semicircular. La neurona motora que comanda los músculos oculares horizontales se activa a una velocidad de , donde es el ángulo de giro horizontal y es su velocidad angular horizontal. Los dos términos dan cuenta de la elasticidad y la viscosidad del tejido ocular. [12]
El momento de inercia rotacional del ojo es insignificante, ya que las personas que usan lentes de contacto ponderados que aumentan el momento de inercia rotacional casi 100 veces aún tienen el mismo VOR (p. 94 [13] ).
El reflejo vestíbulo-ocular debe ser rápido: para una visión clara, el movimiento de la cabeza debe compensarse casi inmediatamente; de lo contrario, la visión corresponde a una fotografía tomada con una mano temblorosa. Las señales se envían desde los canales semicirculares utilizando solo tres neuronas, llamadas arco de tres neuronas . Esto da como resultado movimientos oculares que se retrasan menos de 10 ms respecto del movimiento de la cabeza. [14] El reflejo vestíbulo-ocular es uno de los reflejos más rápidos del cuerpo humano.
Cuando una persona sigue el movimiento de algo con los ojos y la cabeza al mismo tiempo, el VOR es contraproducente para el objetivo de mantener la mirada y el ángulo de la cabeza alineados. Las investigaciones indican que existen mecanismos en el cerebro para suprimir el VOR utilizando la retroalimentación visual activa (retinal) obtenida al observar el objeto en movimiento. [15] En ausencia de retroalimentación visual, como cuando el objeto pasa detrás de una barrera opaca, los humanos pueden continuar rastreando visualmente la posición aparente del objeto utilizando sistemas anticipatorios (extrarretinianos) dentro del cerebro, y el VOR también se suprime durante esta actividad. El VOR incluso puede suprimirse cognitivamente, como cuando se sigue un objetivo imaginario con los ojos y la cabeza al mismo tiempo, aunque el efecto tiende a ser menos dramático que con la retroalimentación visual. [16]
La "ganancia" del VOR se define como el cambio en el ángulo del ojo dividido por el cambio en el ángulo de la cabeza durante el giro de la cabeza. Lo ideal es que la ganancia del VOR rotacional sea 1,0. La ganancia del VOR horizontal y vertical suele ser cercana a 1,0, pero la ganancia del VOR torsional (rotación alrededor de la línea de visión) es generalmente baja. [4] La ganancia del VOR traslacional debe ajustarse a la distancia, debido a la geometría del paralaje de movimiento. Cuando la cabeza se traslada, la dirección angular de los objetivos cercanos cambia más rápido que la dirección angular de los objetivos lejanos. [6]
Si la ganancia del VOR es incorrecta (diferente de 1), por ejemplo, si los músculos oculares están débiles o si una persona se pone un par de anteojos nuevos, el movimiento de la cabeza produce un movimiento de la imagen en la retina, lo que produce visión borrosa. En tales condiciones, el aprendizaje motor ajusta la ganancia del VOR para producir un movimiento ocular más preciso. Esto es lo que se conoce como adaptación del VOR.
Las personas miopes que habitualmente usan gafas negativas tienen una ganancia de RVO menor. Las personas hipermétropes o afágicas que habitualmente usan gafas positivas tienen una ganancia de RVO mayor. Las personas que habitualmente usan lentes de contacto no muestran cambios en la ganancia de RVO. La adaptación monocular y desconjugada del RVO es posible, por ejemplo, después de una parálisis de los músculos extraoculares . (p. 27 [17] )
La fase del VOR también puede adaptarse. [18]
El integrador oculomotor es un integrador con fugas, con un tiempo de fuga característico de ~20 s. Si el tiempo de fuga es demasiado bajo, se produce algún tipo de adaptación para "reparar la fuga" y aumentar el tiempo de fuga. Se plantea la hipótesis de que el integrador con fugas está construido por un circuito de retroalimentación con una ganancia ligeramente inferior a 1, y la adaptación se produce ajustando la ganancia del circuito de retroalimentación. La hipótesis se prueba utilizando un tambor optocinético con un patrón especial que simula el efecto visual de tener un integrador oculomotor con muchas fugas. Después de 1 hora de visualización, el integrador se vuelve "antifugas", lo que significa que su valor crece exponencialmente incluso en ausencia de entrada. El movimiento del ojo se vuelve de retroalimentación positiva, lo que significa que si está ligeramente a la izquierda de un objetivo de fijación, se desviará aún más a la izquierda, y lo mismo para la derecha. También se acompaña de náuseas. [19] (p. 84 [13] )

El consumo de etanol puede alterar el VOR, reduciendo la agudeza visual dinámica. [20] En condiciones normales, la cúpula y la endolinfa tienen la misma densidad (ambas son ). Después de ingerir etanol, el etanol se difunde en la cúpula antes de difundirse en la endolinfa, porque está más cerca de los capilares sanguíneos. Esto hace que la cúpula sea temporalmente más ligera. En este estado, si una persona se recuesta con la mejilla derecha tocando el suelo, entonces la cúpula en la oreja izquierda flotaría hacia la izquierda, creando una sensación ilusoria de rotación lenta de la cabeza de izquierda a derecha. Para compensar esto, el VOR mueve los ojos hacia la izquierda lentamente hasta que llega al límite, y los ojos tiran hacia la derecha rápidamente (nistagmo). Este es el nistagmo posicional del alcohol , fase I (PAN I). La estimulación vestibular inusual también causó síntomas de mareo por movimiento: ilusiones de rotaciones corporales, mareos y náuseas. Estos síntomas desaparecen en unos segundos después de asumir una postura erguida. [21]
Después de un tiempo, la densidad de la cúpula y la endolinfa se iguala, eliminando el efecto del nistagmo. Una vez que el etanol se metaboliza por completo, la cúpula vuelve primero a su densidad normal, lo que crea nistagmo en la dirección opuesta (PAN II) durante la resaca . [21]
Como se predijo, el consumo de agua pesada (1,1 de densidad de agua) tiene el efecto de nistagmo exactamente opuesto en comparación con el consumo de etanol. El consumo de una mezcla de agua pesada ( ) y etanol ( ) anula en gran medida el efecto. [21] La macroglobulinemia , o el consumo de glicerol (1,26 de densidad de agua), tiene efectos similares al agua pesada. [22] [23] [24]
Este reflejo se puede comprobar mediante la prueba de impulso cefálico rápido o prueba de Halmagyi-Curthoys , en la que se mueve rápidamente la cabeza hacia un lado con fuerza y se controla si los ojos consiguen mantener la mirada en la misma dirección. Cuando la función del sistema de equilibrio derecho se ve reducida, ya sea por una enfermedad o por un accidente, ya no se puede percibir correctamente un movimiento rápido de la cabeza hacia la derecha. Como consecuencia, no se genera ningún movimiento ocular compensatorio y el paciente no puede fijar un punto en el espacio durante este movimiento rápido de la cabeza.
La prueba de impulso cefálico se puede realizar junto a la cama y se puede utilizar como herramienta de detección de problemas con el sistema vestibular de una persona. [25] También se puede evaluar con fines diagnósticos mediante una prueba de impulso cefálico por video (VHIT). En esta prueba diagnóstica, una persona usa gafas de alta sensibilidad que detectan cambios rápidos en el movimiento ocular. Esta prueba puede brindar información específica del sitio sobre el sistema vestibular y su función. [26]
Otra forma de comprobar la respuesta del VOR es la prueba del reflejo calórico , que consiste en intentar inducir nistagmo (movimiento compensatorio de los ojos en ausencia de movimiento de la cabeza) vertiendo agua fría o caliente en el oído. También existen irrigaciones calóricas con aire bitérmico, en las que se administra aire caliente y frío en el oído. [27]
El reflejo vestíbulo-ocular puede evaluarse mediante la prueba del reflejo calórico antes mencionada ; ésta desempeña un papel importante en la confirmación del diagnóstico de muerte del tronco encefálico. En este proceso se debe seguir un código de práctica, a saber, el de la Academy of Medical Royal Colleges. [28]
Resumen: El reflejo cervicoocular, también conocido por su acrónimo COR, implica la consecución de la estabilización de un objetivo visual, [29] y de la imagen en la retina, mediante ajustes de la mirada impactados por los movimientos o rotaciones del cuello y/o de la cabeza. El proceso funciona en conjunto con el reflejo vestíbulo-ocular (VOR). [30] Es evidente en ciertos animales que no pueden mover mucho los ojos, como los búhos. [31]






