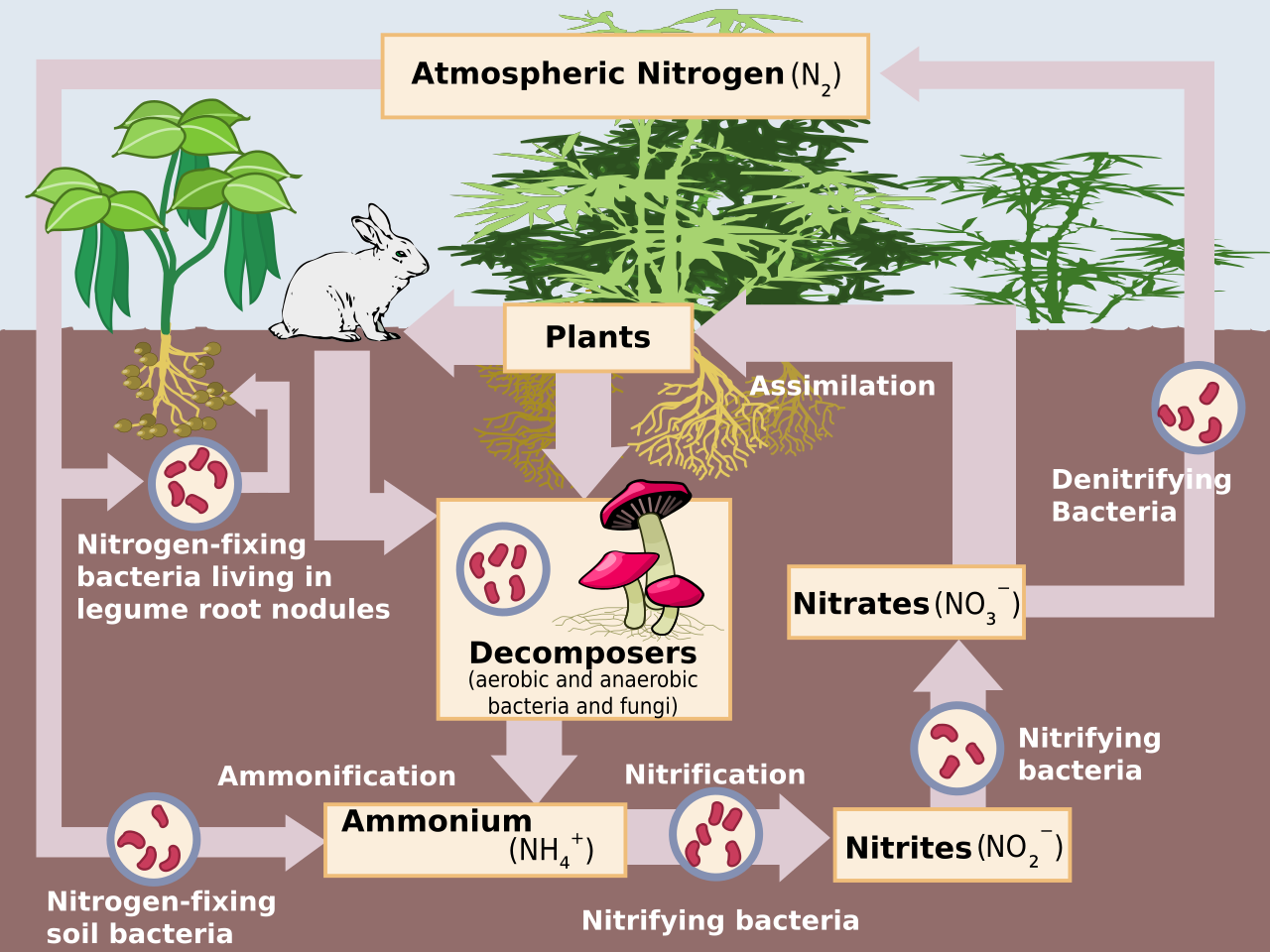
Algunas bacterias fijadoras de nitrógeno tienen relaciones simbióticas con plantas , especialmente legumbres , musgos y helechos acuáticos como la Azolla . [4] Las relaciones no simbióticas más laxas entre diazótrofos y plantas a menudo se denominan asociativas, como se ve en la fijación de nitrógeno en las raíces del arroz . La fijación de nitrógeno ocurre entre algunas termitas y hongos . [5] Ocurre naturalmente en el aire por medio de la producción de NO x por los rayos . [6] [7]
"Las prolongadas investigaciones sobre la relación de las plantas con la adquisición de nitrógeno iniciadas por de Saussure , Ville , Lawes , Gilbert y otros, culminaron con el descubrimiento de la fijación simbiótica por Hellriegel y Wilfarth en 1887". [12]
"Los experimentos de Bossingault en 1855 y de Pugh, Gilbert y Lawes en 1887 habían demostrado que el nitrógeno no entraba directamente en la planta. El descubrimiento del papel de las bacterias fijadoras de nitrógeno por Herman Hellriegel y Herman Wilfarth en 1886-1888 abriría una nueva era en la ciencia del suelo ". [13]
La fijación biológica de nitrógeno (FBN) ocurre cuando el nitrógeno atmosférico se convierte en amoníaco por acción de una enzima nitrogenasa . [1] La reacción general para la FBN es:
El proceso está acoplado a la hidrólisis de 16 equivalentes de ATP y está acompañado de la coformación de un equivalente de H 2. [14] La conversión de N 2La conversión en amoníaco se produce en un grupo metálico llamado FeMoco , una abreviatura del cofactor hierro- molibdeno . El mecanismo se lleva a cabo mediante una serie de pasos de protonación y reducción en los que el sitio activo de FeMoco hidrogena el N 2sustrato. [15] En diazótrofos de vida libre , el amoníaco generado por la nitrogenasa se asimila en glutamato a través de la vía glutamina sintetasa /glutamato sintetasa. Los genes nif microbianos necesarios para la fijación de nitrógeno están ampliamente distribuidos en diversos entornos. [16]
Por ejemplo, se ha demostrado que la madera en descomposición, que generalmente tiene un bajo contenido de nitrógeno, alberga una comunidad diazotrófica. [17] [18] Las bacterias enriquecen el sustrato de madera con nitrógeno a través de la fijación, lo que permite la descomposición de la madera muerta por hongos. [19]
Las nitrogenasas se degradan rápidamente en presencia de oxígeno. Por este motivo, muchas bacterias dejan de producir la enzima en presencia de oxígeno. Muchos organismos fijadores de nitrógeno existen únicamente en condiciones anaeróbicas , respirando para reducir los niveles de oxígeno o uniendo el oxígeno con una proteína como la leghemoglobina . [20] [21]
Importancia del nitrógeno
El nitrógeno atmosférico es inaccesible para la mayoría de los organismos, [22] debido a que su triple enlace covalente es muy fuerte. La mayoría absorbe nitrógeno fijado de diversas fuentes. Por cada 100 átomos de carbono, se asimilan aproximadamente de 2 a 20 átomos de nitrógeno. La relación atómica de carbono (C): nitrógeno (N): fósforo (P) observada en promedio en la biomasa planctónica fue descrita originalmente por Alfred Redfield, [23] quien determinó la relación estequiométrica entre átomos de C:N:P, la relación de Redfield, en 106:16:1. [23]
Nitrogenasa
El complejo proteico nitrogenasa es responsable de catalizar la reducción del gas nitrógeno (N 2 ) a amoniaco (NH 3 ). [24] [25] En las cianobacterias , este sistema enzimático está alojado en una célula especializada llamada heterocisto . [26] La producción del complejo nitrogenasa está regulada genéticamente, y la actividad del complejo proteico depende de las concentraciones ambientales de oxígeno y de las concentraciones intra y extracelulares de amoniaco y especies de nitrógeno oxidado (nitrato y nitrito). [27] [28] [29] Además, se cree que las concentraciones combinadas de amonio y nitrato inhiben la fijación de N , específicamente cuando las concentraciones intracelulares de 2- oxoglutarato (2-OG) superan un umbral crítico. [30] La célula heterocisto especializada es necesaria para el rendimiento de la nitrogenasa como resultado de su sensibilidad al oxígeno ambiental. [31]
La nitrogenasa consta de dos proteínas, una proteína catalítica dependiente del hierro, comúnmente conocida como proteína MoFe y una proteína reductora solo de hierro (proteína Fe). Hay tres proteínas dependientes del hierro diferentes, dependiente del molibdeno , dependiente del vanadio y solo de hierro , y las tres variaciones de la proteína nitrogenasa contienen un componente de proteína de hierro. La nitrogenasa dependiente del molibdeno es la nitrogenasa presente más comúnmente. [32] Los diferentes tipos de nitrogenasa se pueden determinar por el componente de proteína de hierro específico. [33] La nitrogenasa está altamente conservada. La expresión génica a través de la secuenciación de ADN puede distinguir qué complejo proteico está presente en el microorganismo y potencialmente se está expresando. Con mayor frecuencia, el gen nif H se utiliza para identificar la presencia de nitrogenasa dependiente de molibdeno, seguido de las nitrogenasas reductasas estrechamente relacionadas (componente II) vnf H y anf H que representan la nitrogenasa dependiente de vanadio y la nitrogenasa solo de hierro, respectivamente. [34] Al estudiar la ecología y la evolución de las bacterias fijadoras de nitrógeno , el gen nifH es el biomarcador más utilizado. [35] nif H tiene dos genes similares anf H y vnfH que también codifican el componente nitrogenasa reductasa del complejo nitrogenasa. [36]
Evolución de la nitrogenasa
Se cree que la nitrogenasa evolucionó en algún momento entre 1.500 y 2.200 millones de años atrás (Ga), [37] [38] aunque hay algunos datos isotópicos que muestran que la evolución de la nitrogenasa se produjo alrededor de 3.200 millones de años atrás. [39] La nitrogenasa parece haber evolucionado a partir de proteínas similares a la madurasa , aunque actualmente se desconoce la función de la proteína precedente. [40]
La nitrogenasa tiene tres formas diferentes ( Nif, Anf y Vnf ) que corresponden al metal que se encuentra en el sitio activo de la proteína (molibdeno, hierro y vanadio respectivamente). [41] Se cree que las abundancias de metales marinos a lo largo de la línea de tiempo geológica de la Tierra han impulsado la abundancia relativa de qué forma de nitrogenasa era más común. [42] Actualmente, no hay un acuerdo concluyente sobre qué forma de nitrogenasa surgió primero.
Las cianobacterias , comúnmente conocidas como algas verdeazuladas, habitan casi todos los entornos iluminados de la Tierra y desempeñan papeles clave en el ciclo del carbono y el nitrógeno de la biosfera . En general, las cianobacterias pueden utilizar varias fuentes inorgánicas y orgánicas de nitrógeno combinado, como nitrato , nitrito , amonio , urea o algunos aminoácidos . Varias cepas de cianobacterias también son capaces de crecimiento diazotrófico, una capacidad que puede haber estado presente en su último ancestro común en el eón Arcaico . [47] La fijación de nitrógeno no solo ocurre naturalmente en los suelos sino también en los sistemas acuáticos, incluidos los de agua dulce y marinos. [48] [49] De hecho, la cantidad de nitrógeno fijado en el océano es al menos tanta como en la tierra. [50] Se cree que la cianobacteria marina colonial Trichodesmium fija el nitrógeno a tal escala que representa casi la mitad de la fijación de nitrógeno en los sistemas marinos a nivel mundial. [51] Los líquenes de la superficie marina y las bacterias no fotosintéticas pertenecientes a Proteobacteria y Planctomycetes fijan nitrógeno atmosférico significativo. [52] Las especies de cianobacterias fijadoras de nitrógeno en aguas dulces incluyen: Aphanizomenon y Dolichospermum (anteriormente Anabaena). [53] Dichas especies tienen células especializadas llamadas heterocitos , en las que la fijación de nitrógeno ocurre a través de la enzima nitrogenasa. [54] [55]
Algas
Un tipo de orgánulo puede convertir el gas nitrógeno en una forma biológicamente disponible. Este nitroplasto fue descubierto en las algas . [56]
Simbiosis de nódulos radiculares
Familia de las leguminosas
En esta raíz de haba se ven nódulos.
Las plantas que contribuyen a la fijación de nitrógeno incluyen las de la familia de las leguminosas —Fabaceae— con taxones como el kudzu , el trébol , la soja , la alfalfa , el lupino , el cacahuete y el rooibos . [44] Contienen bacterias rizobias simbióticas dentro de los nódulos de sus sistemas radiculares , que producen compuestos nitrogenados que ayudan a la planta a crecer y competir con otras plantas. [57] Cuando la planta muere, el nitrógeno fijado se libera, poniéndolo a disposición de otras plantas; esto ayuda a fertilizar el suelo . [20] [58] La gran mayoría de las leguminosas tienen esta asociación, pero algunos géneros (por ejemplo, Styphnolobium ) no la tienen. En muchas prácticas agrícolas tradicionales, los campos se rotan a través de varios tipos de cultivos, que normalmente incluyen uno que consiste principalmente o totalmente en trébol . [ cita requerida ]
La eficiencia de fijación en el suelo depende de muchos factores, entre ellos la leguminosa y las condiciones del aire y del suelo. Por ejemplo, la fijación de nitrógeno por el trébol rojo puede variar de 50 a 200 lb/acre (56 a 224 kg/ha). [59]
No leguminosas
Un nódulo de raíz de aliso seccionado
La capacidad de fijar nitrógeno en nódulos está presente en plantas actinorícicas como el aliso y el arrayán , con la ayuda de las bacterias Frankia . Se encuentran en 25 géneros en los órdenes Cucurbitales , Fagales y Rosales , que junto con los Fabales forman un clado de fijación de nitrógeno de eurosidas . La capacidad de fijar nitrógeno no está universalmente presente en estas familias. Por ejemplo, de 122 géneros de Rosaceae , solo cuatro fijan nitrógeno. Fabales fue el primer linaje en ramificarse de este clado de fijación de nitrógeno; por lo tanto, la capacidad de fijar nitrógeno puede ser plesiomórfica y posteriormente perdida en la mayoría de los descendientes de la planta fijadora de nitrógeno original; sin embargo, puede ser que los requisitos genéticos y fisiológicos básicos estuvieran presentes en un estado incipiente en los ancestros comunes más recientes de todas estas plantas, pero solo evolucionaron hasta la función completa en algunas de ellas. [60]
Además, Trema ( Parasponia ), un género tropical de la familia Cannabaceae , es excepcionalmente capaz de interactuar con rizobios y formar nódulos fijadores de nitrógeno. [61]
Otros simbiontes vegetales
Algunas otras plantas viven en asociación con un cianobionte (cianobacteria como Nostoc ) que les fija nitrógeno:
En 1784, Henry Cavendish describió por primera vez un método para fijar el nitrógeno mediante arcos eléctricos que hacían reaccionar el nitrógeno y el oxígeno del aire. Este método se implementó en el proceso Birkeland-Eyde de 1903. [66] La fijación del nitrógeno mediante rayos es un proceso natural muy similar.
La posibilidad de que el nitrógeno atmosférico reaccione con ciertas sustancias químicas fue observada por primera vez por Desfosses en 1828. Observó que las mezclas de óxidos de metales alcalinos y carbono reaccionan con el nitrógeno a altas temperaturas. Con el uso de carbonato de bario como material de partida, el primer proceso comercial estuvo disponible en la década de 1860, desarrollado por Margueritte y Sourdeval. El cianuro de bario resultante reacciona con vapor, produciendo amoníaco. En 1898, Frank y Caro desarrollaron lo que se conoce como el proceso Frank-Caro para fijar el nitrógeno en forma de cianamida de calcio . El proceso fue eclipsado por el proceso Haber , que se descubrió en 1909. [67] [68]
Proceso de Haber
Equipo para el estudio de la fijación de nitrógeno por rayos alfa (Laboratorio de Investigación de Nitrógeno Fijo, 1926)
El método industrial dominante para producir amoníaco es el proceso Haber, también conocido como proceso Haber-Bosch. [69] La producción de fertilizantes es ahora la mayor fuente de nitrógeno fijo producido por el hombre en el ecosistema terrestre . El amoníaco es un precursor necesario para fertilizantes , explosivos y otros productos. El proceso Haber requiere altas presiones (alrededor de 200 atm) y altas temperaturas (al menos 400 °C), que son condiciones rutinarias para la catálisis industrial. Este proceso utiliza gas natural como fuente de hidrógeno y aire como fuente de nitrógeno. El producto de amoníaco ha dado lugar a una intensificación de los fertilizantes nitrogenados a nivel mundial [70] y se le atribuye el apoyo a la expansión de la población humana de alrededor de 2 mil millones a principios del siglo XX a aproximadamente 8 mil millones de personas en la actualidad. [71]
Catálisis homogénea
Se han llevado a cabo muchas investigaciones para descubrir catalizadores para la fijación de nitrógeno, a menudo con el objetivo de reducir los requisitos de energía. Sin embargo, hasta ahora, dichas investigaciones no han logrado acercarse a la eficiencia y facilidad del proceso Haber. Muchos compuestos reaccionan con el nitrógeno atmosférico para dar complejos de dinitrógeno . El primer complejo de dinitrógeno del que se informó fue Ru(NH 3) 5( Nota 2) 2+ . [72] Algunos complejos solubles catalizan la fijación de nitrógeno. [73]
El nitrógeno se puede fijar mediante un rayo que convierte el gas nitrógeno ( N 2) y gas oxígeno ( O 2) en la atmósfera en NO x ( óxidos de nitrógeno ). El N 2La molécula es altamente estable y no reactiva debido al triple enlace entre los átomos de nitrógeno. [74] Los rayos producen suficiente energía y calor para romper este enlace [74], lo que permite que los átomos de nitrógeno reaccionen con el oxígeno y formen NO. incógnitaEstos compuestos no pueden ser utilizados por las plantas, pero a medida que esta molécula se enfría, reacciona con el oxígeno para formar NO. 2, [75] que a su vez reacciona con agua para producir HNO 2( ácido nitroso ) o HNO 3( ácido nítrico ). Cuando estos ácidos se filtran en el suelo, forman NO 3 - (nitrato) , que es útil para las plantas. [76] [74]
^ ab Howard JB, Rees DC (1996). "Base estructural de la fijación biológica del nitrógeno". Chemical Reviews . 96 (7): 2965–2982. doi :10.1021/cr9500545. PMID 11848848.
^ Burris RH, Wilson PW (junio de 1945). "Fijación biológica del nitrógeno". Revista anual de bioquímica . 14 (1): 685–708. doi :10.1146/annurev.bi.14.070145.003345. ISSN 0066-4154.
^ Wagner SC (2011). «Biological Nitrogen Fixation». Nature Education Knowledge . 3 (10): 15. Archivado desde el original el 13 de septiembre de 2018. Consultado el 29 de enero de 2019 .
^ Zahran HH (diciembre de 1999). "Simbiosis rizobio-leguminosa y fijación de nitrógeno en condiciones severas y en un clima árido". Microbiology and Molecular Biology Reviews . 63 (4): 968–89, tabla de contenidos. doi :10.1128/MMBR.63.4.968-989.1999. PMC 98982 . PMID 10585971.
^ Sapountzis P, de Verges J, Rousk K, Cilliers M, Vorster BJ, Poulsen M (2016). "Potencial de fijación de nitrógeno en la simbiosis entre hongos y termitas". Frontiers in Microbiology . 7 : 1993. doi : 10.3389/fmicb.2016.01993 . PMC 5156715 . PMID 28018322.
^ Slosson E (1919). Química creativa. Nueva York, NY: The Century Co. págs. 19–37.
^ Hill RD, Rinker RG, Wilson HD (1979). "Fijación atmosférica de nitrógeno por rayos". J. Atmos. Sci . 37 (1): 179–192. Código Bibliográfico :1980JAtS...37..179H. doi : 10.1175/1520-0469(1980)037<0179:ANFBL>2.0.CO;2 .
^ Boussingault (1838). "Recherches chimiques sur la vegetarian, entreprises dans le but d'examiner si les plantes prennent de l'azote à l'atmosphere" [Investigaciones químicas sobre la vegetación, realizadas con el objetivo de examinar si las plantas absorben nitrógeno en la atmósfera]. Annales de Chimie et de Physique . 2da serie (en francés). 67 : 5–54.y 69: 353–367.
^ Smil V (2001). Enriqueciendo la Tierra . Instituto Tecnológico de Massachusetts.
^ Hellriegel H, Wilfarth H (1888). Untersuchungen über die Stickstoffnahrung der Gramineen und Leguminosen [ Estudios sobre la ingesta de nitrógeno de Gramineae y Leguminosae ] (en alemán). Berlín, Alemania: Buchdruckerei der "Post" Kayssler & Co.
^ Beijerinck MW (1901). "Über oligonitrophile Mikroben" [Sobre microbios oligonitrófilos]. Centralblatt für Bakteriologie, Parasitenkunde, Infektionskrankheiten und Hygiene (en alemán). 7 (16): 561–582.
^ Howard S. Reed (1942) Una breve historia de la ciencia vegetal , página 230, Chronic Publishing
^ Lee CC, Ribbe MW, Hu Y (2014). Kroneck PM, Sosa Torres ME (eds.). "Capítulo 7. Escisión del triple enlace N,N: la transformación de dinitrógeno en amoníaco por nitrogenasas". Iones metálicos en las ciencias de la vida . 14 . Springer: 147–76. doi :10.1007/978-94-017-9269-1_7. PMID 25416394.
^ Hoffman BM, Lukoyanov D, Dean DR, Seefeldt LC (febrero de 2013). "Nitrogenasa: un mecanismo preliminar". Accounts of Chemical Research . 46 (2): 587–95. doi :10.1021/ar300267m. PMC 3578145 . PMID 23289741.
^ Gaby JC, Buckley DH (julio de 2011). "Un censo global de la diversidad de la nitrogenasa". Microbiología ambiental . 13 (7): 1790–9. Bibcode :2011EnvMi..13.1790G. doi :10.1111/j.1462-2920.2011.02488.x. PMID 21535343.
^ Rinne KT, Rajala T, Peltoniemi K, Chen J, Smolander A, Mäkipää R (2017). "Tasas de acumulación y fuentes de nitrógeno externo en madera en descomposición en un bosque dominado por la pícea de Noruega". Ecología funcional . 31 (2): 530–541. Bibcode :2017FuEco..31..530R. doi : 10.1111/1365-2435.12734 . ISSN 1365-2435. S2CID 88551895.
^ Hoppe B, Kahl T, Karasch P, Wubet T, Bauhus J, Buscot F, et al. (2014). "El análisis de redes revela vínculos ecológicos entre las bacterias fijadoras de nitrógeno y los hongos que descomponen la madera". PLOS ONE . 9 (2): e88141. Bibcode :2014PLoSO...988141H. doi : 10.1371/journal.pone.0088141 . PMC 3914916 . PMID 24505405.
^ Tláskal V, Brabcová V, Větrovský T, Jomura M, López-Mondéjar R, Oliveira Monteiro LM, et al. (enero de 2021). "Las funciones complementarias de los hongos y bacterias que habitan en la madera facilitan la descomposición de la madera muerta". mSistemas . 6 (1). doi :10.1128/mSystems.01078-20. PMC 7901482 . PMID 33436515.
^ ab Postgate J (1998). Fijación de nitrógeno (3.ª ed.). Cambridge: Cambridge University Press.
^ Streicher SL, Gurney EG, Valentine RC (octubre de 1972). "Los genes de fijación del nitrógeno". Nature . 239 (5374): 495–9. Bibcode :1972Natur.239..495S. doi :10.1038/239495a0. PMID 4563018. S2CID 4225250.
^ Delwiche CC (1983). "Ciclado de elementos en la biosfera". En Läuchli A, Bieleski RL (eds.). Nutrición vegetal inorgánica . Enciclopedia de fisiología vegetal. Berlín, Heidelberg: Springer. págs. 212–238. doi :10.1007/978-3-642-68885-0_8. ISBN978-3-642-68885-0.
^ ab Redfield AC (1958). "El control biológico de los factores químicos en el medio ambiente". Científico estadounidense . 46 (3): 230A–221. ISSN 0003-0996. JSTOR 27827150.
^ Seefeldt LC, Yang ZY, Lukoyanov DA, Harris DF, Dean DR, Raugei S, et al. (2020). "Reducción de sustratos por nitrogenasas". Chemical Reviews . 120 (12): 5082–5106. doi :10.1021/acs.chemrev.9b00556. PMC 7703680 . PMID 32176472.
^ Threatt SD, Rees DC (2023). "Fijación biológica de nitrógeno en teoría, práctica y realidad: una perspectiva sobre el sistema de la nitrogenasa del molibdeno". FEBS Letters . 597 (1): 45–58. doi :10.1002/1873-3468.14534. PMC 10100503 . PMID 36344435.
^ Peterson RB, Wolk CP (diciembre de 1978). "Alta recuperación de la actividad de la nitrogenasa y de la nitrogenasa marcada con Fe en heterocistos aislados de Anabaena variabilis". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 75 (12): 6271–6275. Bibcode :1978PNAS...75.6271P. doi : 10.1073/pnas.75.12.6271 . PMC 393163 . PMID 16592599.
^ Beversdorf LJ, Miller TR, McMahon KD (6 de febrero de 2013). "El papel de la fijación de nitrógeno en la toxicidad de la floración de cianobacterias en un lago templado y eutrófico". PLOS ONE . 8 (2): e56103. Bibcode :2013PLoSO...856103B. doi : 10.1371/journal.pone.0056103 . PMC 3566065 . PMID 23405255.
^ Gallon JR (1 de marzo de 2001). "Fijación de N2 en fotótrofos: adaptación a una forma de vida especializada". Planta y suelo . 230 (1): 39–48. doi :10.1023/A:1004640219659. ISSN 1573-5036. S2CID 22893775.
^ Paerl H (9 de marzo de 2017). "La paradoja de la fijación de nitrógeno por cianobacterias en aguas naturales". F1000Research . 6 : 244. doi : 10.12688/f1000research.10603.1 . PMC 5345769 . PMID 28357051.
^ Li JH, Laurent S, Konde V, Bédu S, Zhang CC (noviembre de 2003). "Un aumento en el nivel de 2-oxoglutarato promueve el desarrollo de heterocistos en la cepa PCC 7120 de la cianobacteria Anabaena sp". Microbiología . 149 (Pt 11): 3257–3263. doi : 10.1099/mic.0.26462-0 . PMID 14600238.
^ Wolk CP, Ernst A, Elhai J (1994). "Metabolismo y desarrollo de heterocistos". En Bryant DA (ed.). La biología molecular de las cianobacterias . Avances en la fotosíntesis. Dordrecht: Springer Netherlands. págs. 769–823. doi :10.1007/978-94-011-0227-8_27. ISBN978-94-011-0227-8.
^ Burgess BK, Lowe DJ (noviembre de 1996). "Mecanismo de la nitrogenasa del molibdeno". Chemical Reviews . 96 (7): 2983–3012. doi :10.1021/cr950055x. PMID 11848849.
^ Schneider K, Müller A (2004). "Nitrogenasa de solo hierro: características catalíticas, estructurales y espectroscópicas excepcionales". En Smith BE, Richards RL, Newton WE (eds.). Catalizadores para la fijación del nitrógeno . Fijación del nitrógeno: orígenes, aplicaciones y progreso de la investigación. Dordrecht: Springer Netherlands. págs. 281–307. doi :10.1007/978-1-4020-3611-8_11. ISBN978-1-4020-3611-8.
^ Knoche KL, Aoyama E, Hasan K, Minteer SD (2017). "Función de la nitrogenasa y la ferredoxina en el mecanismo de fijación bioelectrocatalítica del nitrógeno por la cianobacteria Anabaena variabilis SA-1 mutante inmovilizada en electrodos de óxido de indio y estaño (ITO)". Electrochimica Acta (en coreano). 232 : 396–403. doi :10.1016/j.electacta.2017.02.148.
^ Raymond J, Siefert JL, Staples CR, Blankenship RE (marzo de 2004). "La historia natural de la fijación del nitrógeno". Biología molecular y evolución . 21 (3): 541–554. doi : 10.1093/molbev/msh047 . PMID 14694078.
^ Schüddekopf K, Hennecke S, Liese U, Kutsche M, Klipp W (mayo de 1993). "Caracterización de genes anf específicos para la nitrogenasa alternativa e identificación de genes nif necesarios para ambas nitrogenasas en Rhodobacter capsulatus". Microbiología molecular . 8 (4): 673–684. doi :10.1111/j.1365-2958.1993.tb01611.x. PMID 8332060. S2CID 42057860.
^ Garcia AK, McShea H, Kolaczkowski B, Kaçar B (mayo de 2020). "Reconstrucción de la historia evolutiva de las nitrogenasas: evidencia de la utilización ancestral del cofactor molibdeno". Geobiología . 18 (3): 394–411. Bibcode :2020Gbio...18..394G. doi :10.1111/gbi.12381. ISSN 1472-4677. PMC 7216921 . PMID 32065506.
^ Boyd ES, Anbar AD, Miller S, Hamilton TL, Lavin M, Peters JW (mayo de 2011). "Un origen metanógeno tardío para la nitrogenasa dependiente del molibdeno". Geobiología . 9 (3): 221–232. Bibcode :2011Gbio....9..221B. doi :10.1111/j.1472-4669.2011.00278.x. ISSN 1472-4677. PMID 21504537.
^ Stüeken EE, Buick R, Guy BM, Koehler MC (abril de 2015). "Evidencia isotópica de fijación biológica de nitrógeno por molibdeno-nitrogenasa de 3,2 mil millones de años". Nature . 520 (7549): 666–669. Bibcode :2015Natur.520..666S. doi :10.1038/nature14180. ISSN 0028-0836. PMID 25686600.
^ Garcia AK, Kolaczkowski B, Kaçar B (2 de marzo de 2022). Archibald J (ed.). "La reconstrucción de los predecesores de la nitrogenasa sugiere su origen a partir de proteínas similares a la madurasa". Genome Biology and Evolution . 14 (3). doi :10.1093/gbe/evac031. ISSN 1759-6653. PMC 8890362 . PMID 35179578.
^ Eady RR (1 de enero de 1996). "Relaciones estructura-función de las nitrogenasas alternativas". Chemical Reviews . 96 (7): 3013–3030. doi :10.1021/cr950057h. ISSN 0009-2665. PMID 11848850.
^ Anbar AD, Knoll AH (16 de agosto de 2002). "Química y evolución del océano proterozoico: ¿un puente bioinorgánico?". Science . 297 (5584): 1137–1142. Bibcode :2002Sci...297.1137A. doi :10.1126/science.1069651. ISSN 0036-8075. PMID 12183619.
^ Instituto MP (6 de agosto de 2021). "Aportes de nitrógeno en el océano antiguo: bacterias poco apreciadas pasan a primer plano".
^ ab Mus F, Crook MB, Garcia K, Garcia Costas A, Geddes BA, Kouri ED, et al. (julio de 2016). Kelly RM (ed.). "Fijación simbiótica de nitrógeno y los desafíos para su extensión a las no leguminosas". Applied and Environmental Microbiology . 82 (13): 3698–3710. Bibcode :2016ApEnM..82.3698M. doi :10.1128/AEM.01055-16. PMC 4907175 . PMID 27084023.
^ Dhamad AE, Lessner DJ (octubre de 2020). Atomi H (ed.). "Un sistema CRISPRi-dCas9 para arqueas y su uso para examinar la función génica durante la fijación de nitrógeno por Methanosarcina acetivorans". Microbiología aplicada y ambiental . 86 (21): e01402–20. Código Bibliográfico :2020ApEnM..86E1402D. doi :10.1128/AEM.01402-20. PMC 7580536 . PMID 32826220.
^ Bae HS, Morrison E, Chanton JP, Ogram A (abril de 2018). "Los metanógenos son los principales contribuyentes a la fijación de nitrógeno en los suelos de los Everglades de Florida". Microbiología aplicada y ambiental . 84 (7): e02222–17. Bibcode :2018ApEnM..84E2222B. doi :10.1128/AEM.02222-17. PMC 5861825 . PMID 29374038.
^ Latysheva N, Junker VL, Palmer WJ, Codd GA, Barker D (marzo de 2012). "La evolución de la fijación de nitrógeno en cianobacterias". Bioinformática . 28 (5): 603–606. doi : 10.1093/bioinformatics/bts008 . PMID 22238262.
^ Pierella Karlusich JJ, Pelletier E, Lombard F, Carsique M, Dvorak E, Colin S, et al. (julio de 2021). "Patrones de distribución global de fijadores de nitrógeno marinos mediante métodos moleculares y de imágenes". Nature Communications . 12 (1): 4160. Bibcode :2021NatCo..12.4160P. doi :10.1038/s41467-021-24299-y. PMC 8260585 . PMID 34230473.
^ Ash C (13 de agosto de 2021). Ash C, Smith J (eds.). "Un poco de luz sobre los diazótrofos". Science . 373 (6556): 755.7–756. Bibcode :2021Sci...373..755A. doi :10.1126/science.373.6556.755-g. ISSN 0036-8075. S2CID 238709371.
^ Kuypers MM, Marchant HK, Kartal B (mayo de 2018). "La red de ciclo del nitrógeno microbiano". Nature Reviews. Microbiología . 16 (5): 263–276. doi :10.1038/nrmicro.2018.9. hdl : 21.11116/0000-0003-B828-1 . PMID 29398704. S2CID 3948918.
^ Bergman B, Sandh G, Lin S, Larsson J, Carpenter EJ (mayo de 2013). "Trichodesmium: una cianobacteria marina muy extendida con propiedades inusuales de fijación de nitrógeno". FEMS Microbiology Reviews . 37 (3): 286–302. doi :10.1111/j.1574-6976.2012.00352.x. PMC 3655545 . PMID 22928644.
^ "Estudio a gran escala indica nuevos y abundantes microbios fijadores de nitrógeno en la superficie del océano". ScienceDaily . Archivado desde el original el 8 de junio de 2019 . Consultado el 8 de junio de 2019 .
^ Rolff C, Almesjö L, Elmgren R (5 de marzo de 2007). "Fijación de nitrógeno y abundancia de la cianobacteria diazotrófica Aphanizomenon sp. en el Báltico propiamente dicho". Marine Ecology Progress Series . 332 : 107–118. Bibcode :2007MEPS..332..107R. doi : 10.3354/meps332107 .
^ Carmichael WW (12 de octubre de 2001). "Efectos sobre la salud de las cianobacterias productoras de toxinas: las CyanoHAB"". Evaluación de riesgos humanos y ecológicos . 7 (5): 1393–1407. Bibcode :2001HERA....7.1393C. doi :10.1080/20018091095087. ISSN 1080-7039. S2CID 83939897.
^ Bothe H, Schmitz O, Yates MG, Newton WE (diciembre de 2010). "Fijación de nitrógeno y metabolismo del hidrógeno en cianobacterias". Microbiology and Molecular Biology Reviews . 74 (4): 529–551. doi :10.1128/MMBR.00033-10. PMC 3008169 . PMID 21119016.
^ Wong C (11 de abril de 2024). "Los científicos descubren las primeras algas que pueden fijar nitrógeno gracias a una estructura celular diminuta". Nature . doi :10.1038/d41586-024-01046-z. PMID 38605201.
^ Kuypers MM, Marchant HK, Kartal B (mayo de 2018). "La red de ciclo del nitrógeno microbiano". Nature Reviews. Microbiología . 16 (5): 263–276. doi :10.1038/nrmicro.2018.9. hdl : 21.11116/0000-0003-B828-1 . PMID 29398704. S2CID 3948918.
^ Smil V (2000). Ciclos de la vida . Biblioteca Scientific American.
^ "Fijación de nitrógeno e inoculación de leguminosas forrajeras" (PDF) . Archivado desde el original (PDF) el 2 de diciembre de 2016.
^ Dawson JO (2008). "Ecología de plantas actinorícicas". Simbiosis actinorícicas fijadoras de nitrógeno . Fijación de nitrógeno: orígenes, aplicaciones y avances de la investigación. Vol. 6. Springer. págs. 199–234. doi :10.1007/978-1-4020-3547-0_8. ISBN .978-1-4020-3540-1.
^ Op den Camp R, Streng A, De Mita S, Cao Q, Polone E, Liu W, et al. (febrero de 2011). "Receptor de micorrizas tipo LysM reclutado para la simbiosis de rizobios en Parasponia no leguminosa". Ciencia . 331 (6019): 909–12. Código Bib : 2011 Ciencia... 331..909O. doi : 10.1126/ciencia.1198181. PMID 21205637. S2CID 20501765.
^ "Biología de las cícadas, artículo 1: Raíces corraloides de las cícadas". www1.biologie.uni-hamburg.de . Consultado el 14 de octubre de 2021 .
^ Rai AN (2000). "Simbiosis entre cianobacterias y plantas". New Phytologist . 147 (3): 449–481. doi : 10.1046/j.1469-8137.2000.00720.x . PMID 33862930.
^ Van Deynze A, Zamora P, Delaux PM, Heitmann C, Jayaraman D, Rajasekar S, et al. (Agosto de 2018). "La fijación de nitrógeno en una variedad local de maíz está respaldada por una microbiota diazotrófica asociada al mucílago". Más biología . 16 (8): e2006352. doi : 10.1371/journal.pbio.2006352 . PMC 6080747 . PMID 30086128.
^ Pskowski M (16 de julio de 2019). "Maíz indígena: ¿Quién posee los derechos sobre la planta 'maravillosa' de México?". Yale E360 .
^ Eyde S (1909). "La fabricación de nitratos a partir de la atmósfera mediante el arco eléctrico: proceso Birkeland-Eyde". Revista de la Royal Society of Arts . 57 (2949): 568–576. JSTOR 41338647.
^ Heinrich H, Nevbner R (1934). "Die Umwandlungsgleichung Ba(CN)2 → BaCN2 + C im Temperaturgebiet von 500 bis 1000 °C" [La reacción de conversión Ba(CN) 2 → BaCN 2 + C en el rango de temperatura de 500 a 1000 °C]. Z. Elektrochem. Angew. Phys. Chem . 40 (10): 693–698. doi :10.1002/bbpc.19340401005. S2CID 179115181. Archivado desde el original el 20 de agosto de 2016 . Consultado el 8 de agosto de 2016 .
^ Curtis HA (1932). Nitrógeno fijo.
^ Smil, V. 2004. Enriqueciendo la Tierra: Fritz Haber, Carl Bosch y la transformación de la producción mundial de alimentos, MIT Press.
^ Gilbert PM , Maranger R , Sobota DJ, Bouwman L (1 de octubre de 2014). "El vínculo entre Haber Bosch y la floración de algas nocivas (HB–HAB)". Environmental Research Letters . 9 (10): 105001. Bibcode :2014ERL.....9j5001G. doi : 10.1088/1748-9326/9/10/105001 . ISSN 1748-9326. S2CID 154724892.
^ Erisman JW, Sutton MA, Galloway J, Klimont Z, Winiwarter W (octubre de 2008). "Cómo un siglo de síntesis de amoníaco cambió el mundo". Nature Geoscience . 1 (10): 636–639. Bibcode :2008NatGe...1..636E. doi :10.1038/ngeo325. ISSN 1752-0908. S2CID 94880859.
^ Allen AD, Senoff CV (1965). "Complejos de nitrógenopentamminrutenio (II)". J. química. Soc., Química. Comunitario. (24): 621–622. doi :10.1039/C19650000621.
^ Chalkley MJ, Drover MW, Peters JC (junio de 2020). "Conversión catalítica de N2 a NH3 (o -N2H4) mediante complejos de coordinación molecular bien definidos". Chemical Reviews . 120 (12): 5582–5636. doi :10.1021/acs.chemrev.9b00638. PMC 7493999 . PMID 32352271.
^ abc Tuck AF (octubre de 1976). "Producción de óxidos de nitrógeno por descargas de rayos". Quarterly Journal of the Royal Meteorological Society . 102 (434): 749–755. Bibcode :1976QJRMS.102..749T. doi :10.1002/qj.49710243404. ISSN 0035-9009.
^ Hill RD (agosto de 1979). "Fijación de nitrógeno atmosférico por rayos". Revista de ciencias atmosféricas . 37 : 179–192. Código Bibliográfico :1980JAtS...37..179H. doi : 10.1175/1520-0469(1980)037<0179:ANFBL>2.0.CO;2 . ISSN 1520-0469.
^ Levin JS (1984). "Fuentes troposféricas de NOx: rayos y biología" . Consultado el 29 de noviembre de 2018 .
"Colección de fotografías del laboratorio de investigación de nitrógeno fijo de Travis P. Hignett // Colecciones digitales del Science History Institute". digital.sciencehistory.org . Consultado el 16 de agosto de 2019 . Colecciones digitales del Instituto de Historia de la Ciencia (fotografías que representan numerosas etapas del proceso de fijación de nitrógeno y los diversos equipos y aparatos utilizados en la producción de nitrógeno atmosférico, incluidos generadores, compresores, filtros, termostatos y hornos de vacío y altos hornos).
"Proceso propuesto para la fijación del nitrógeno atmosférico", perspectiva histórica, Scientific American , 13 de julio de 1878, pág. 21
Una instantánea global de los fijadores de nitrógeno del océano mediante la comparación de secuencias con células en el océano Tara



.jpg/1280px-Lightning_Pritzerbe_01_(MK).jpg)