La osteopontina ( OPN ), también conocida como sialoproteína ósea I ( BSP-1 o BNSP ), activación temprana de linfocitos T ( ETA-1 ), fosfoproteína secretada 1 ( SPP1 ), 2ar y resistencia a Rickettsia ( Ric ), [5] es una proteína que en los humanos está codificada por el gen SPP1 (fosfoproteína secretada 1). El ortólogo murino es Spp1 . La osteopontina es una SIBLING ( glicoproteína ) que se identificó por primera vez en 1986 en osteoblastos .
El prefijo osteo- indica que la proteína se expresa en el hueso , aunque también se expresa en otros tejidos. El sufijo -pontin se deriva de "pons", la palabra latina para puente, y significa el papel de la osteopontina como proteína de enlace. La osteopontina es una proteína estructural extracelular y, por lo tanto, un componente orgánico del hueso .
OPN es una proteína de matriz extracelular altamente cargada negativamente, fuertemente fosforilada que carece de una estructura secundaria extensa como una proteína intrínsecamente desordenada. [6] [7] Está compuesta por alrededor de 300 aminoácidos (297 en ratón; 314 en humano) y se expresa como una proteína naciente de 33 kDa; también hay sitios de escisión funcionalmente importantes. OPN puede pasar por modificaciones postraduccionales , que aumentan su peso molecular aparente a aproximadamente 44 kDa. [8] El gen OPN está compuesto por 7 exones, 6 de los cuales contienen secuencia codificante. [9] [10] Los primeros dos exones contienen la región 5' no traducida (5' UTR). [11] Los exones 2, 3, 4, 5, 6 y 7 codifican 17, 13, 27, 14, 108 y 134 aminoácidos, respectivamente. [11] Todos los límites intrón-exón son del tipo fase 0, por lo que el empalme alternativo de exones mantiene el marco de lectura del gen OPN.
Figura 1. Sitios de escisión proteolítica para la osteopontina de longitud completa (OPN-FL). La trombina expone el epítopo escindido SVVYGLR (OPN-R) y luego la CPB elimina la arginina c-terminal de OPN-R. El epítopo escindido tiene un dominio no RGD, que se une a los receptores de integrina (α4β1, α9β1 y α9β4). Junto al epítopo escindido, hay un dominio RGD que interactúa con otros receptores de integrina (αvβ1,3,5 y α5β1). No se muestra aquí la gran cantidad de sitios de escisión a lo largo de toda la longitud de la proteína, tal como se degrada por la enzima PHEX expresada por las células de tejido mineralizado. [12]
Isoformas
La OPN de longitud completa (OPN-FL) puede modificarse mediante escisión de trombina , que expone una secuencia críptica, SVVYGLR en la forma escindida de la proteína conocida como OPN-R (Fig. 1). Esta OPN escindida por trombina (OPN-R) expone un epítopo para los receptores de integrina de α4β1, α9β1 y α9β4. [13] [14] Estos receptores de integrina están presentes en varias células inmunes como mastocitos , [15] neutrófilos, [16] y células T. También se expresa en monocitos y macrófagos. [17] Al unirse a estos receptores, las células utilizan varias vías de transducción de señales para provocar respuestas inmunes en estas células. La OPN-R puede escindirse aún más mediante la carboxipeptidasa B (CPB) mediante la eliminación de la arginina C-terminal y convertirse en OPN-L. La función de OPN-L es en gran parte desconocida.
Parece que una variante intracelular de OPN (iOPN) está involucrada en varios procesos celulares, incluida la migración, la fusión y la motilidad. [18] [19] [20] [21] La OPN intracelular se genera utilizando un sitio de inicio de traducción alternativo en la misma especie de ARNm utilizada para generar la isoforma extracelular. [22] Este sitio de inicio de traducción alternativo se encuentra aguas abajo de la secuencia de señal de direccionamiento al retículo endoplásmico N-terminal, lo que permite la traducción citoplasmática de OPN.
Se ha observado que varios cánceres humanos, incluido el cáncer de mama, expresan variantes de empalme de OPN. [23] [24] Las variantes de empalme específicas del cáncer son osteopontina-a, osteopontina-b y osteopontina-c. El exón 5 falta en la osteopontina-b, mientras que la osteopontina-c carece del exón 4. [23] Se ha sugerido que la osteopontina-c facilita el fenotipo independiente del anclaje de algunas células de cáncer de mama humano debido a su incapacidad para asociarse con la matriz extracelular. [23]
Distribución de tejidos
Osteopontina observada en una muestra de tejido pulmonar de un paciente con fibrosis pulmonar idiopática.
La regulación de la expresión del gen de la osteopontina no se comprende completamente. Los diferentes tipos de células pueden diferir en sus mecanismos reguladores del gen OPN. La expresión de OPN en el hueso ocurre predominantemente por osteoblastos y osteocitos (células formadoras de hueso), así como osteoclastos (células reabsortivas del hueso). [29] Los factores de transcripción Runx2 (también conocido como Cbfa1) y osterix (Osx) son necesarios para la expresión de OPN [30] Runx2 y Osx se unen a los promotores de genes específicos de osteoblastos como Col1α1 , Bsp y Opn y regulan positivamente la transcripción. [31]
La hipocalcemia y la hipofosfatemia (casos que estimulan a las células del túbulo proximal del riñón a producir calcitriol (1α,25-dihidroxivitamina D3)) conducen a aumentos en la transcripción, traducción y secreción de OPN. [32] Esto se debe a la presencia de un elemento de respuesta a la vitamina D de alta especificidad ( VDRE ) en el promotor del gen OPN. [33] [34] [35]
La expresión de osteopontina (OPN) está modulada por el antígeno del huevo de Schistosoma mansoni . [36]
Los antígenos de los huevos de Schistosoma mansoni estimulan directamente la expresión de la molécula profibrogénica osteopontina (OPN), y los niveles sistémicos de OPN se correlacionan fuertemente con la gravedad de la enfermedad, lo que sugiere su uso como un posible biomarcador de morbilidad. La investigación sobre el impacto del uso de Praziquantel en los niveles sistémicos de OPN y en la deposición de colágeno hepático en la esquistosomiasis murina crónica reveló que el tratamiento con Praziquantel redujo significativamente los niveles sistémicos de OPN y la deposición de colágeno hepático, lo que indica que la OPN podría ser una herramienta confiable para monitorear la eficacia de PZQ y la regresión de la fibrosis. [37] [36]
El fosfato inorgánico extracelular (ePi) también se ha identificado como un modulador de la expresión de OPN. [38]
La estimulación de la expresión de OPN también ocurre tras la exposición de las células a citocinas proinflamatorias, [39] mediadores clásicos de la inflamación aguda (por ejemplo, factor de necrosis tumoral α [TNFα], interleucina-1β [IL-1β]), angiotensina II, factor de crecimiento transformante β (TGFβ) y hormona paratiroidea (PTH), [40] [41] aunque todavía no se conoce un entendimiento detallado del mecanismo de estas vías reguladoras. También se sabe que la hiperglucemia y la hipoxia aumentan la expresión de OPN. [40] [42] [43]
Función
Apoptosis
La OPN es un factor antiapoptótico importante en muchas circunstancias. La OPN bloquea la muerte celular inducida por activación de los macrófagos y las células T, así como de los fibroblastos y las células endoteliales expuestas a estímulos nocivos. [44] [45] La OPN previene la muerte celular no programada en la colitis inflamatoria. [46]
Biomineralización
OPN pertenece a una familia de proteínas ácidas secretadas (SIBLINGs, Small Integrin Binding LIgand N-Glycosylated proteinas) cuyos miembros tienen una abundancia de aminoácidos cargados negativamente como Asp y Glu . [47] OPN también tiene una gran cantidad de sitios de secuencia de consenso para la fosforilación postraduccional de residuos Ser para formar fosfoserina , proporcionando carga negativa adicional. [48] Se han identificado tramos contiguos de alta carga negativa en OPN y se han denominado motivo poliAsp (ácido poliaspártico) y motivo ASARM (motivo ácido rico en serina y aspartato), y esta última secuencia tiene múltiples sitios de fosforilación. [49] [50] [51] [52] Esta carga negativa general de OPN, junto con sus motivos ácidos específicos y el hecho de que OPN es una proteína intrínsecamente desordenada [53] [6] que permite estructuras abiertas y flexibles, permiten que OPN se una fuertemente a los átomos de calcio disponibles en las superficies de los cristales en varios biominerales. [52] [54] [55] Tal unión de OPN a varios tipos de biominerales basados en calcio ‒como el mineral de fosfato de calcio en huesos y dientes, [56] el mineral de carbonato de calcio en otoconias del oído interno [57] y cáscaras de huevo de aves, [58] y el mineral de oxalato de calcio en cálculos renales [59] [60] [61] — actúa como un inhibidor de la mineralización al estabilizar las fases precursoras minerales transitorias y al unirse directamente a las superficies de los cristales, todo lo cual regula el crecimiento de los cristales. [62] [63] [64]
La OPN es una proteína sustrato para varias enzimas cuyas acciones pueden modular la función inhibidora de la mineralización de la OPN. PHEX (homólogo de la endopeptidasa reguladora de fosfato ligado al cromosoma X) es una de esas enzimas, que degrada ampliamente la OPN y cuyas mutaciones genéticas inactivadoras (en la hipofosfatemia ligada al cromosoma X, XLH) conducen a un procesamiento alterado de la OPN de modo que la OPN inhibidora no puede degradarse y se acumula en la matriz extracelular del hueso (y del diente), lo que contribuye localmente a la osteomalacia (huesos blandos hipomineralizados y odontomalacia (dientes blandos) característica de la XLH. [65] [66] [12] Una relación que describe la regulación local, fisiológica doble negativa (inhibición de inhibidores) de la mineralización que involucra a OPN se ha denominado Principio de Esténcil de la mineralización, por el cual los pares enzima-sustrato imprimen patrones de mineralización en la matriz extracelular (descrito más notablemente para el hueso) degradando inhibidores de la mineralización ( por ejemplo , la enzima TNAP que degrada la inhibición del pirofosfato y la enzima PHEX que degrada la inhibición de la osteopontina). [63] [64] En relación con las enfermedades de mineralización, el Principio de Esténcil es particularmente relevante para la osteomalacia y odontomalacia observadas en la hipofosfatasia y la hipofosfatemia ligada al cromosoma X.
Además de su papel en la regulación de la mineralización normal dentro de las matrices extracelulares de los huesos y los dientes, [67] la OPN también se regula positivamente en sitios de calcificación ectópica patológica [68] [69] —como por ejemplo, en la urolitiasis [59] [61] y la calcificación vascular [70] [68] —, presumiblemente al menos en parte para inhibir la mineralización debilitante en estos tejidos blandos.
Remodelación ósea
Osteoclasto
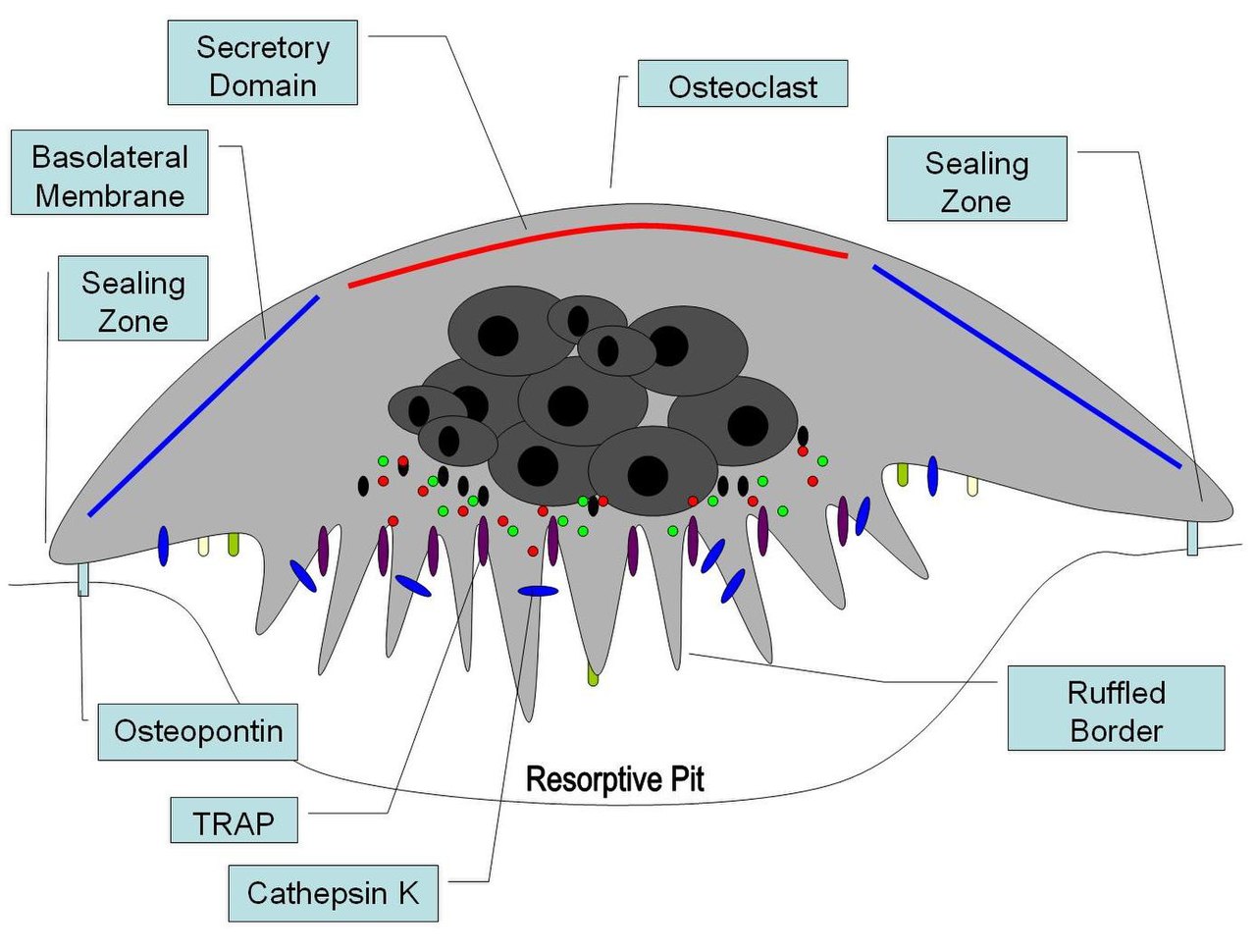
La osteopontina se ha implicado como un factor importante en la remodelación ósea . [71] Específicamente, OPN ancla osteoclastos a la superficie de los huesos donde se inmoviliza por sus propiedades de unión a minerales permitiendo el uso posterior de su motivo RGD para la unión de la integrina de osteoclastos para la fijación y migración celular. [15] OPN en superficies óseas se encuentra en una capa orgánica delgada, la llamada lámina limitans. [72] La parte orgánica del hueso es aproximadamente el 20% del peso seco y cuenta, además de osteopontina, colágeno tipo I , osteocalcina , osteonectina y fosfatasa alcalina . El colágeno tipo I cuenta para el 90% de la masa proteica. La parte inorgánica del hueso es el mineral hidroxiapatita , Ca 10 (PO 4 ) 6 (OH) 2 . La pérdida de hueso puede conducir a osteoporosis , ya que el hueso se agota de calcio si este no se suministra en la dieta.
La OPN sirve para iniciar el proceso mediante el cual los osteoclastos desarrollan sus bordes ondulados para comenzar la resorción ósea. La OPN contiene un motivo de unión a la integrina RGD.
Activación celular
Las células T activadas son promovidas por IL-12 para diferenciarse hacia el tipo Th1, produciendo citocinas que incluyen IL-12 e IFNγ . OPN inhibe la producción de la citocina Th2 IL-10, lo que conduce a una respuesta Th1 mejorada. OPN influye en la inmunidad mediada por células y tiene funciones de citocina Th1. Mejora la producción y proliferación de inmunoglobulina de células B. [7] OPN también induce la desgranulación de mastocitos. [73] La anafilaxia mediada por IgE se reduce significativamente en ratones knock-out de OPN en comparación con ratones de tipo salvaje. El papel de OPN en la activación de macrófagos también se ha implicado en un cáncer ya que los tumores productores de OPN fueron capaces de inducir la activación de macrófagos en comparación con los tumores deficientes en OPN. [74]
Fig. 2. Funciones inmunológicas conocidas de la OPN. La OPN se une a varios receptores de integrina, incluidos α4β1, α9β1 y α9β4 expresados por leucocitos, y se sabe que inducen la adhesión celular, la migración y la supervivencia en células inmunes, incluidos neutrófilos, macrófagos, células T, mastocitos y osteoclastos.
Quimiotaxis
La OPN desempeña un papel importante en el reclutamiento de neutrófilos en la enfermedad hepática alcohólica. [16] [75] La OPN es importante para la migración de neutrófilos in vitro . [76] Además, la OPN recluta células inflamatorias a las articulaciones artríticas en el modelo de artritis inducida por colágeno de la artritis reumatoide. [77] [78] Un estudio in vitro reciente en 2008 ha descubierto que la OPN desempeña un papel en la migración de mastocitos. [73] Aquí se cultivaron mastocitos knock out de OPN y observaron un nivel disminuido de quimiotaxis en estas células en comparación con los mastocitos de tipo salvaje. También se descubrió que la OPN actúa como un factor quimiotáctico de macrófagos. [79] En el mono rhesus, la OPN evita que los macrófagos abandonen el sitio de acumulación en los cerebros, lo que indica un mayor nivel de quimiotaxis.
Sistema inmunitario
La OPN se une a varios receptores de integrina, incluidos α4β1, α9β1 y α9β4 expresados por los leucocitos . Se ha demostrado que estos receptores funcionan en la adhesión celular, la migración y la supervivencia en estas células.
La osteopontina (OPN) se expresa en una variedad de células inmunes, incluidos macrófagos, neutrófilos , células dendríticas, microglia y células T y B , con una cinética variable. Se informa que la OPN actúa como un modulador inmunológico de diversas maneras. [7] En primer lugar, tiene propiedades quimiotácticas , que promueven el reclutamiento de células a sitios inflamatorios. También funciona como una proteína de adhesión , involucrada en la unión celular y la cicatrización de heridas . Además, la OPN media la activación celular y la producción de citocinas , además de promover la supervivencia celular regulando la apoptosis . [7] Se encuentran los siguientes ejemplos. [7]
Importancia clínica
El hecho de que la OPN interactúe con múltiples receptores de la superficie celular que se expresan de forma ubicua la convierte en un actor activo en muchos procesos fisiológicos y patológicos, entre los que se incluyen la cicatrización de heridas, el recambio óseo, la tumorigénesis, la inflamación, la isquemia y las respuestas inmunitarias. La manipulación de los niveles plasmáticos (o locales) de OPN puede ser útil en el tratamiento de enfermedades autoinmunes, metástasis del cáncer, enfermedades de mineralización ósea (y dental), osteoporosis y algunas formas de estrés. [7]
Enfermedades autoinmunes
La OPN se ha visto implicada en la patogénesis de la artritis reumatoide . La OPN-R, la forma de OPN escindida por trombina, se encuentra elevada en las articulaciones afectadas por artritis reumatoide. Sin embargo, el papel de la OPN en la artritis reumatoide aún no está claro. Un grupo descubrió que los ratones knock out de OPN estaban protegidos contra la artritis. [80] mientras que otros no pudieron reproducir esta observación. [81]
Se ha descubierto que la OPN desempeña un papel en otras enfermedades autoinmunes, incluidas la hepatitis autoinmune , la enfermedad alérgica de las vías respiratorias y la esclerosis múltiple . [82]
Alergia y asma
Recientemente se ha asociado la osteopontina con la inflamación alérgica y el asma . La expresión de Opn aumenta significativamente en las células epiteliales y subepiteliales pulmonares de pacientes asmáticos en comparación con sujetos sanos. [83] La expresión de Opn también aumenta en los pulmones de ratones con inflamación alérgica de las vías respiratorias. [83] La forma secretada de Opn (Opn-s) desempeña un papel proinflamatorio durante la sensibilización al alérgeno (OVA/Alum), ya que la neutralización de Opn-s durante esa fase da como resultado una inflamación alérgica de las vías respiratorias significativamente más leve. [83] Por el contrario, la neutralización de Opn-s durante el desafío antigénico exacerba la enfermedad alérgica de las vías respiratorias. [83] Estos efectos de Opn-s están mediados principalmente por la regulación de las células dendríticas plasmocitoides (DC) supresoras de Th2 durante la sensibilización primaria y las DC convencionales promotoras de Th2 durante el desafío antigénico secundario. [83] También se informó que la deficiencia de OPN protege contra la remodelación y la hiperreactividad bronquial (BHR), nuevamente utilizando un modelo de desafío crónico con alérgenos de remodelación de las vías respiratorias. [84] Además, recientemente se demostró que la expresión de OPN está regulada positivamente en el asma humano, está asociada con cambios de remodelación y su expresión subepitelial se correlaciona con la gravedad de la enfermedad. [85] También se ha informado que la OPN aumenta en el sobrenadante de esputo de asmáticos fumadores, [86] así como en el BALF y el tejido bronquial de controles fumadores y asmáticos. [87]
Colitis
La expresión de Opn aumenta en la enfermedad inflamatoria intestinal (EII). [88] La expresión de Opn aumenta en gran medida en las células intestinales inmunes y no inmunes y en el plasma de pacientes con enfermedad de Crohn (EC) y colitis ulcerosa (CU), así como en el colon y el plasma de ratones con colitis experimental. [88] [89] [90] Los niveles plasmáticos aumentados de Opn están relacionados con la gravedad de la inflamación de la EC, y ciertos haplotipos del gen Opn (Spp1) son modificadores de la susceptibilidad a la EC. Opn también tiene un papel proinflamatorio en la colitis inducida por TNBS y sulfato de dextrano sódico (DSS), que son modelos murinos para la EII. Se encontró que Opn estaba altamente expresada por un subconjunto específico de células dendríticas (DC) derivadas de ganglios linfáticos mesentéricos murinos (MLN) y es altamente proinflamatoria para la colitis. [91] Las células dendríticas son importantes para el desarrollo de la inflamación intestinal en humanos con EII y en ratones con colitis experimental. La expresión de Opn por este subconjunto de DC MLN inflamatorio es crucial para su acción patogénica durante la colitis. [91]
La osteopontina está implicada en la progresión de la enfermedad PDAC ( adenocarcinoma pancreático ). [93] Se expresa como una de las tres variantes de empalme en PDAC, con osteopontina-a expresada en casi todos los PDAC, la expresión de osteopontina-b correlacionada con la supervivencia y la osteopontina-c correlacionada con la enfermedad metastásica. Debido a que PDAC secreta formas de osteopontina empalmadas alternativamente, muestra potencial para la focalización específica del tumor y la etapa de la enfermedad. Aunque se desconocen los mecanismos exactos de la señalización de osteopontina en PDAC, se une a CD44 e integrinas para desencadenar procesos como la progresión tumoral y la inhibición del complemento. La osteopontina también impulsa la metástasis al desencadenar la liberación del factor de crecimiento endotelial vascular (VEGF) y la metaloproteasa de matriz (MMP), que se inhibe al derribar la osteopontina. Este proceso también es estimulado por la nicotina, que es el mecanismo propuesto por el cual los fumadores experimentan un riesgo elevado de PC. La osteopontina se está explorando como un marcador para PC. Se ha descubierto que su rendimiento es mejor que el de CA19.9 para distinguir entre IPMN [80] y PDAC resecable de pancreatitis. Se están desarrollando anticuerpos antiosteopontina, incluido hu1A12, que inhibió la metástasis en un estudio in vivo y también cuando se hibridó con el anticuerpo anti-VEGF bevacizumab. Al menos un ensayo clínico está explorando el uso de osteopontina como marcador de hipoxia intratumoral. Sin embargo, este marcador sigue siendo relativamente inexplorado. [94]
La osteopontina también está implicada en la formación excesiva de cicatrices y se ha desarrollado un gel para inhibir su efecto. [95]
Aunque Opn promueve la metástasis y puede usarse como un biomarcador del cáncer, los últimos estudios describieron nuevas funciones protectoras de la molécula en poblaciones de células innatas durante el desarrollo del tumor. En particular, el mantenimiento de un grupo de células asesinas naturales (NK) con una función inmunológica óptima es crucial para la defensa del huésped contra la formación de tumores cancerosos. Un estudio en PNAS describe que iOpn es un componente molecular esencial responsable del mantenimiento de la expansión funcional de las células NK. La ausencia de iOPN da como resultado la incapacidad de mantener la celularidad normal de las NK y el aumento de la muerte celular después de la estimulación por la citocina IL-15. Las células NK deficientes en OPN no logran navegar con éxito la fase de contracción de la respuesta inmunológica, lo que resulta en una expansión deficiente de las células NK de larga vida y respuestas defectuosas a las células tumorales. [98]
Además, las células dendríticas plasmocitoides (pDC) protegen del melanoma, y este efecto está mediado por los IFN de tipo I. [99] Un estudio publicado en JCB demostró que un fragmento específico (SLAYGLR) de la proteína Opn puede hacer que las pDC sean más “aptas” para proteger contra el desarrollo del melanoma. Esto se logró mediante la activación de un nuevo eje integrina α4/IFN-β que es independiente de MyD88 y opera a través de una vía PI3K/mTOR/IRF3. [100]
Insuficiencia cardiaca
La osteopontina se expresa mínimamente en condiciones normales, pero se acumula rápidamente a medida que disminuye la función cardíaca. [101] [102] Específicamente, desempeña un papel central en la respuesta de remodelación al infarto de miocardio , y se regula positivamente de manera drástica en la miocardiopatía hipertrófica (HCM) y dilatada (DCM) . [102] Una vez abundante, estimula una amplia gama de cambios fisiológicos en el miocardio , incluida la angiogénesis , la producción local de citocinas , la diferenciación de miofibroblastos , el aumento de la deposición de la matriz extracelular y la hipertrofia de los cardiomiocitos . En conjunto, estos procesos remodelan la estructura del corazón, reduciendo en efecto su capacidad para funcionar normalmente y aumentando el riesgo de insuficiencia cardíaca . [103] [104]
Enfermedad de Parkinson
La OPN desempeña un papel en el estrés oxidativo y nitrosativo, la apoptosis, la disfunción mitocondrial y la excitotoxicidad, que también están implicadas en la patogénesis de la enfermedad de Parkinson . Se estudiaron las concentraciones de OPN en suero y líquido cefalorraquídeo (LCR) de pacientes con EP, lo que demuestra que los niveles de OPN en el líquido corporal están elevados en pacientes con EP. [105]
Enfermedades y lesiones musculares
Se están acumulando evidencias que sugieren que la osteopontina desempeña una serie de funciones en enfermedades del músculo esquelético , como la distrofia muscular de Duchenne . La osteopontina se ha descrito como un componente del entorno inflamatorio de los músculos distróficos y lesionados, [28] [106] [107] [108] y también se ha demostrado que aumenta la cicatrización de los músculos del diafragma de ratones distróficos envejecidos. [109] Un estudio reciente ha identificado la osteopontina como un determinante de la gravedad de la enfermedad en pacientes con distrofia muscular de Duchenne . [110] Este estudio encontró que una mutación en el promotor del gen de la osteopontina, conocido por causar niveles bajos de expresión de osteopontina, está asociada con una disminución de la edad hasta la pérdida de la deambulación y la fuerza muscular en pacientes con distrofia muscular de Duchenne .
Artrosis de cadera
Se ha observado un aumento de los niveles plasmáticos de OPN en pacientes con artrosis idiopática de cadera. Además, se ha observado una correlación entre los niveles plasmáticos de OPN y la gravedad de la enfermedad. [111]
Implantación de óvulos fecundados
La OPN se expresa en las células endometriales durante la implantación. Debido a la producción de progesterona por los ovarios, la OPN se regula al alza enormemente para ayudar en este proceso. El endometrio debe sufrir decidualización, el proceso en el que el endometrio sufre cambios para prepararse para la implantación, lo que conducirá a la adhesión del embrión. El endometrio alberga células del estroma que se diferenciarán para producir un entorno óptimo para que el embrión se adhiera (decidualización). La OPN es una proteína vital para la proliferación y diferenciación de las células del estroma, y se une al receptor αvβ3 para ayudar con la adhesión. La OPN junto con la decidualización en última instancia fomenta la implantación exitosa del embrión temprano. Una inactivación del gen OPN da como resultado inestabilidad de la adhesión en la interfaz materno-fetal. [112] [113]
Referencias
^ abc GRCh38: Lanzamiento de Ensembl 89: ENSG00000118785 – Ensembl , mayo de 2017
^ abc GRCm38: Lanzamiento de Ensembl 89: ENSMUSG00000029304 – Ensembl , mayo de 2017
^ "Referencia de PubMed humana:". Centro Nacional de Información Biotecnológica, Biblioteca Nacional de Medicina de EE. UU .
^ "Referencia de PubMed sobre ratón". Centro Nacional de Información Biotecnológica, Biblioteca Nacional de Medicina de EE. UU .
^ "Gen Entrez: fosfoproteína 1 secretada por SPP1".
^ ab Kalmar L, Homola D, Varga G, Tompa P (septiembre de 2012). "El desorden estructural en las proteínas aporta orden al crecimiento de los cristales en la biomineralización". Bone . 51 (3): 528–534. doi :10.1016/j.bone.2012.05.009. PMID 22634174.
^ abcdefg Wang KX, Denhardt DT (2008). "Osteopontina: papel en la regulación inmunitaria y las respuestas al estrés". Cytokine & Growth Factor Reviews . 19 (5–6): 333–345. doi :10.1016/j.cytogfr.2008.08.001. PMID 18952487.
^ Rangaswami H, Bulbule A, Kundu GC (febrero de 2006). "Osteopontina: papel en la señalización celular y la progresión del cáncer". Tendencias en biología celular . 16 (2): 79–87. doi :10.1016/j.tcb.2005.12.005. PMID 16406521.
^ Young MF, Kerr JM, Termine JD, Wewer UM, Wang MG, McBride OW, Fisher LW (agosto de 1990). "Clonación de ADNc, distribución y heterogeneidad del ARNm, localización cromosómica y análisis RFLP de la osteopontina humana (OPN)". Genómica . 7 (4): 491–502. doi :10.1016/0888-7543(90)90191-V. PMID 1974876.
^ Kiefer MC, Bauer DM, Barr PJ (abril de 1989). "El ADNc y la secuencia de aminoácidos derivada de la osteopontina humana". Nucleic Acids Research . 17 (8): 3306. doi :10.1093/nar/17.8.3306. PMC 317745 . PMID 2726470.
^ ab Crosby AH, Edwards SJ, Murray JC, Dixon MJ (mayo de 1995). "Organización genómica del gen de la osteopontina humana: exclusión del locus de un papel causal en la patogénesis de la dentinogénesis imperfecta tipo II". Genómica . 27 (1): 155–160. doi :10.1006/geno.1995.1018. PMID 7665163.
^ ab Barros NM, Hoac B, Neves RL, Addison WN, Assis DM, Murshed M, et al. (marzo de 2013). "Procesamiento proteolítico de osteopontina por PHEX y acumulación de fragmentos de osteopontina en el hueso del ratón Hyp, el modelo murino de hipofosfatemia ligada al cromosoma X". Journal of Bone and Mineral Research . 28 (3): 688–699. doi : 10.1002/jbmr.1766 . PMID 22991293. S2CID 20840491.
^ Laffón A, García-Vicuña R, Humbría A, Postigo AA, Corbí AL, de Landázuri MO, Sánchez-Madrid F (agosto de 1991). "Expresión y función reguladas al alza de los receptores de fibronectina VLA-4 en células T humanas activadas en la artritis reumatoide". La Revista de Investigación Clínica . 88 (2): 546–552. doi :10.1172/JCI115338. PMC 295383 . PMID 1830891.
^ Seiffge D (diciembre de 1996). "Efectos protectores del anticuerpo monoclonal contra VLA-4 sobre la adhesión leucocitaria y el curso de la enfermedad en la artritis adyuvante en ratas". The Journal of Rheumatology . 23 (12): 2086–91. PMID 8970045.
^ ab Reinholt FP, Hultenby K, Oldberg A, Heinegård D (junio de 1990). "Osteopontina: un posible anclaje de los osteoclastos al hueso". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 87 (12): 4473–5. Bibcode :1990PNAS...87.4473R. doi : 10.1073/pnas.87.12.4473 . PMC 54137 . PMID 1693772.
^ ab Banerjee A, Apte UM, Smith R, Ramaiah SK (marzo de 2006). "La mayor infiltración de neutrófilos mediada por osteopontina es un factor que probablemente contribuye a la mayor susceptibilidad de las mujeres a la enfermedad hepática alcohólica". The Journal of Pathology . 208 (4): 473–485. doi : 10.1002/path.1917 . PMID 16440289. S2CID 45879742.
^ Sodek J, Batista Da Silva AP, Zohar R (mayo de 2006). "Osteopontina y protección de la mucosa". Revista de investigación dental . 85 (5): 404–415. doi :10.1177/154405910608500503. PMID 16632752. S2CID 25898774.[ enlace muerto permanente ]
^ Zohar R, Suzuki N, Suzuki K, Arora P, Glogauer M, McCulloch CA, Sodek J (julio de 2000). "La osteopontina intracelular es un componente integral del complejo CD44-ERM involucrado en la migración celular". Journal of Cellular Physiology . 184 (1): 118–130. doi :10.1002/(SICI)1097-4652(200007)184:1<118::AID-JCP13>3.0.CO;2-Y. PMID 10825241. S2CID 11548419.
^ Suzuki K, Zhu B, Rittling SR, Denhardt DT, Goldberg HA, McCulloch CA, Sodek J (agosto de 2002). "La colocalización de osteopontina intracelular con CD44 está asociada con la migración, la fusión celular y la resorción en osteoclastos". Journal of Bone and Mineral Research . 17 (8): 1486–97. doi :10.1359/jbmr.2002.17.8.1486. PMID 12162503. S2CID 21307088.
^ Zhu B, Suzuki K, Goldberg HA, Rittling SR, Denhardt DT, McCulloch CA, Sodek J (enero de 2004). "La osteopontina modula la quimiotaxis dependiente de CD44 de los macrófagos peritoneales a través de receptores acoplados a proteína G: evidencia de un papel para una forma intracelular de osteopontina". Journal of Cellular Physiology . 198 (1): 155–167. doi :10.1002/jcp.10394. PMID 14584055. S2CID 21075107.
^ Junaid A, Moon MC, Harding GE, Zahradka P (febrero de 2007). "La osteopontina se localiza en el núcleo de las células 293 y se asocia con la quinasa tipo polo-1". American Journal of Physiology. Fisiología celular . 292 (2): C919–C926. doi :10.1152/ajpcell.00477.2006. PMID 17005603.
^ Shinohara ML, Kim HJ, Kim JH, Garcia VA, Cantor H (mayo de 2008). "La traducción alternativa de osteopontina genera isoformas intracelulares y secretadas que median distintas actividades biológicas en células dendríticas". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 105 (20): 7235–9. Bibcode :2008PNAS..105.7235S. doi : 10.1073/pnas.0802301105 . PMC 2438233 . PMID 18480255.
^ abc He B, Mirza M, Weber GF (abril de 2006). "Una variante de empalme de osteopontina induce independencia de anclaje en células de cáncer de mama humano". Oncogene . 25 (15): 2192–2202. doi :10.1038/sj.onc.1209248. PMID 16288209. S2CID 6632395.
^ Mirza M, Shaughnessy E, Hurley JK, Vanpatten KA, Pestano GA, He B, Weber GF (febrero de 2008). "La osteopontina-c es un marcador selectivo del cáncer de mama". Revista internacional del cáncer . 122 (4): 889–897. doi : 10.1002/ijc.23204 . PMID 17960616. S2CID 22146646.
^ Ashizawa N, Graf K, Do YS, Nunohiro T, Giachelli CM, Meehan WP, et al. (noviembre de 1996). "La osteopontina es producida por fibroblastos cardíacos de rata y media la síntesis de ADN inducida por A(II) y la contracción del gel de colágeno". The Journal of Clinical Investigation . 98 (10): 2218–27. doi :10.1172/JCI119031. PMC 507670 . PMID 8941637.
^ Murry CE, Giachelli CM, Schwartz SM, Vracko R (diciembre de 1994). "Los macrófagos expresan osteopontina durante la reparación de la necrosis miocárdica". The American Journal of Pathology . 145 (6): 1450–62. PMC 1887495 . PMID 7992848.
^ Ikeda T, Shirasawa T, Esaki Y, Yoshiki S, Hirokawa K (diciembre de 1993). "El ARNm de osteopontina se expresa en células espumosas derivadas del músculo liso en lesiones ateroscleróticas humanas de la aorta". The Journal of Clinical Investigation . 92 (6): 2814–20. doi :10.1172/JCI116901. PMC 288482 . PMID 8254036.
^ ab Uaesoontrachoon K, Yoo HJ, Tudor EM, Pike RN, Mackie EJ, Pagel CN (abril de 2008). "Osteopontina y mioblastos del músculo esquelético: asociación con la regeneración muscular y regulación de la función de los mioblastos in vitro". The International Journal of Biochemistry & Cell Biology . 40 (10): 2303–14. doi :10.1016/j.biocel.2008.03.020. PMID 18490187.
^ Merry K, Dodds R, Littlewood A, Gowen M (abril de 1993). "Expresión del ARNm de osteopontina por osteoclastos y osteoblastos en el modelado de hueso humano adulto". Journal of Cell Science . 104 (4): 1013–20. doi :10.1242/jcs.104.4.1013. PMID 8314886.
^ Nakashima K, Zhou X, Kunkel G, Zhang Z, Deng JM, Behringer RR, de Crombrugghe B (enero de 2002). "El nuevo factor de transcripción que contiene dedos de zinc osterix es necesario para la diferenciación de osteoblastos y la formación ósea". Cell . 108 (1): 17–29. doi : 10.1016/S0092-8674(01)00622-5 . PMID 11792318. S2CID 14030684.
^ Ducy P, Zhang R, Geoffroy V, Ridall AL, Karsenty G (mayo de 1997). "Osf2/Cbfa1: un activador transcripcional de la diferenciación de osteoblastos". Cell . 89 (5): 747–754. doi : 10.1016/S0092-8674(00)80257-3 . PMID 9182762. S2CID 16979723.
^ Yucha C, Guthrie D (diciembre de 2003). "Homeostasis renal del calcio". Nephrology Nursing Journal . 30 (6): 755–764. PMID 14730782.
^ Prince CW, Butler WT (septiembre de 1987). "La 1,25-dihidroxivitamina D3 regula la biosíntesis de osteopontina, una proteína de unión celular derivada del hueso, en células de osteosarcoma similares a osteoblastos clonales". Collagen and Related Research . 7 (4): 305–313. doi :10.1016/s0174-173x(87)80036-5. PMID 3478171.
^ Oldberg A, Jirskog-Hed B, Axelsson S, Heinegård D (diciembre de 1989). "Regulación del ARNm de la sialoproteína ósea por hormonas esteroides". The Journal of Cell Biology . 109 (6 Pt 1): 3183–6. doi :10.1083/jcb.109.6.3183. PMC 2115918 . PMID 2592421.
^ Chang PL, Prince CW (abril de 1991). "La 1 alfa,25-dihidroxivitamina D3 estimula la síntesis y secreción de osteopontina no fosforilada (fosfoproteína 1 secretada) en células epidérmicas JB6 de ratón". Cancer Research . 51 (8): 2144–50. PMID 2009532.
^ ab Pereira TA, Vaz de Melo Trindade G, Trindade Santos E, Pereira FE, Souza MM (mayo de 2021). "La farmacoterapia con praziquantel reduce los niveles de osteopontina sistémica y el contenido de colágeno hepático en la esquistosomiasis mansoni murina". Revista Internacional de Parasitología . 51 (6): 437–440. doi :10.1016/j.ijpara.2020.11.002. PMID 33493521. S2CID 231711719.
^ Gill HS, LeJambre LF (1996). "Prefacio: nuevos enfoques para el control de los parásitos helmintos del ganado. Actas de una conferencia internacional. Armidale, Nueva Gales del Sur, 18-21 de abril de 1995". Int J Parasitol . 26 (8–9): 797–8. doi :10.1016/S0020-7519(96)80042-7. PMID 8923128.
^ Fatherazi S, Matsa-Dunn D, Foster BL, Rutherford RB, Somerman MJ, Presland RB (enero de 2009). "El fosfato regula la transcripción del gen de la osteopontina". Revista de investigación dental . 88 (1): 39–44. doi :10.1177/0022034508328072. PMC 3128439 . PMID 19131315.
^ Guo H, Cai CQ, Schroeder RA, Kuo PC (enero de 2001). "La osteopontina es un regulador de retroalimentación negativa de la síntesis de óxido nítrico en macrófagos murinos". Journal of Immunology . 166 (2): 1079–86. doi : 10.4049/jimmunol.166.2.1079 . PMID 11145688.
^ ab Ricardo SD, Franzoni DF, Roesener CD, Crisman JM, Diamond JR (mayo de 2000). "Inhibición antisentido de la traducción de osteopontina por angiotensinógeno y AT(1) en células tubulares proximales de rata". American Journal of Physiology. Renal Physiology . 278 (5): F708–F716. doi :10.1152/ajprenal.2000.278.5.F708. PMID 10807582.
^ Noda M, Rodan GA (febrero de 1989). "Regulación transcripcional de la producción de osteopontina en células similares a osteoblastos de rata por la hormona paratiroidea". The Journal of Cell Biology . 108 (2): 713–8. doi :10.1083/jcb.108.2.713. PMC 2115413 . PMID 2465299.
^ Hullinger TG, Pan Q, Viswanathan HL, Somerman MJ (enero de 2001). "Activación del promotor OPN por TGFbeta y BMP-2: funciones de los elementos de unión a smad y hox". Experimental Cell Research . 262 (1): 69–74. doi :10.1006/excr.2000.5074. PMID 11120606.
^ Sodhi CP, Phadke SA, Batlle D, Sahai A (abril de 2001). "La hipoxia y la glucosa alta provocan un crecimiento exagerado de las células mesangiales y la síntesis de colágeno: papel de la osteopontina". Revista estadounidense de fisiología. Fisiología renal . 280 (4): F667–F674. doi :10.1152/ajprenal.2001.280.4.F667. PMID 11249858. S2CID 26145407.
^ Denhardt DT, Noda M, O'Regan AW, Pavlin D, Berman JS (mayo de 2001). "La osteopontina como un medio para hacer frente a las agresiones ambientales: regulación de la inflamación, remodelación tisular y supervivencia celular". The Journal of Clinical Investigation . 107 (9): 1055–61. doi :10.1172/JCI12980. PMC 209291 . PMID 11342566.
^ Standal T, Borset M, Sundan A (septiembre de 2004). "El papel de la osteopontina en la adhesión, la migración, la supervivencia celular y la remodelación ósea". Oncología experimental . 26 (3): 179–184. PMID 15494684.
^ Da Silva AP, Pollett A, Rittling SR, Denhardt DT, Sodek J, Zohar R (septiembre de 2006). "La destrucción tisular exacerbada en la colitis aguda inducida por DSS de ratones sin OPN se asocia con la regulación negativa de la expresión de TNF-alfa y la muerte celular no programada". Journal of Cellular Physiology . 208 (3): 629–639. doi :10.1002/jcp.20701. PMID 16741956. S2CID 33704508.
^ Fisher LW, Fedarko NS (2003). "Seis genes expresados en huesos y dientes codifican los miembros actuales de la familia de proteínas SIBLING". Connective Tissue Research . 44 (Supl 1): 33–40. doi :10.1080/03008200390152061. PMID 12952171. S2CID 29910722.
^ Christensen B, Nielsen MS, Haselmann KF, Petersen TE, Sørensen ES (agosto de 2005). "Los residuos modificados postraduccionalmente de la osteopontina humana nativa se encuentran en grupos: identificación de 36 sitios de fosforilación y cinco de O-glicosilación y sus implicaciones biológicas". The Biochemical Journal . 390 (Pt 1): 285–292. doi :10.1042/BJ20050341. PMC 1184582 . PMID 15869464.
^ David V, Martin A, Hedge AM, Drezner MK, Rowe PS (marzo de 2011). "Péptidos ASARM: regulación del fosfato sérico dependiente e independiente de PHEX". Revista estadounidense de fisiología. Fisiología renal . 300 (3): F783–F791. doi :10.1152/ajprenal.00304.2010. PMC 3064126. PMID 21177780 .
^ Martin A, David V, Laurence JS, Schwarz PM, Lafer EM, Hedge AM, Rowe PS (abril de 2008). "Degradación de MEPE, DMP1 y liberación de péptidos ASARM HERMANOS (minhibinas): los péptidos ASARM son directamente responsables de la mineralización defectuosa en HYP". Endocrinología . 149 (4): 1757–72. doi :10.1210/en.2007-1205. PMC 2276704 . PMID 18162525.
^ Addison WN, Nakano Y, Loisel T, Crine P, McKee MD (octubre de 2008). "Los péptidos MEPE-ASARM controlan la mineralización de la matriz extracelular mediante la unión a la hidroxiapatita: una inhibición regulada por la escisión de ASARM por PHEX". Journal of Bone and Mineral Research . 23 (10): 1638–49. doi : 10.1359/jbmr.080601 . PMID 18597632. S2CID 7709872.
^ ab Addison WN, Masica DL, Gray JJ, McKee MD (abril de 2010). "La inhibición dependiente de la fosforilación de la mineralización por péptidos ASARM de osteopontina está regulada por la escisión de PHEX". Journal of Bone and Mineral Research . 25 (4): 695–705. doi :10.1359/jbmr.090832. PMID 19775205. S2CID 25689595.
^ Kurzbach D, Platzer G, Schwarz TC, Henen MA, Konrat R, Hinderberger D (agosto de 2013). "Despliegue cooperativo de conformaciones compactas de la proteína intrínsecamente desordenada osteopontina". Bioquímica . 52 (31): 5167–75. doi :10.1021/bi400502c. PMC 3737600 . PMID 23848319.
^ Azzopardi PV, O'Young J, Lajoie G, Karttunen M, Goldberg HA, Hunter GK (febrero de 2010). "Funciones de la electrostática y la conformación en las interacciones proteína-cristal". PLOS ONE . 5 (2): e9330. Bibcode :2010PLoSO...5.9330A. doi : 10.1371/journal.pone.0009330 . PMC 2824833 . PMID 20174473.
^ Hunter GK, O'Young J, Grohe B, Karttunen M, Goldberg HA (diciembre de 2010). "La hipótesis del polielectrolito flexible de la interacción proteína-biomineral". Langmuir . 26 (24): 18639–46. doi :10.1021/la100401r. PMID 20527831.
^ McKee MD, Nanci A (mayo de 1995). "Inmunocitoquímica de oro coloidal postincrustación de proteínas de la matriz extracelular no colágena en tejidos mineralizados". Microscopy Research and Technique . 31 (1): 44–62. doi :10.1002/jemt.1070310105. PMID 7626799. S2CID 5767310.
^ Takemura T, Sakagami M, Nakase T, Kubo T, Kitamura Y, Nomura S (septiembre de 1994). "Localización de osteopontina en los órganos otoconiales de ratas adultas". Hearing Research . 79 (1–2): 99–104. doi :10.1016/0378-5955(94)90131-7. PMID 7806488. S2CID 4765287.
^ Hincke MT, Nys Y, Gautron J, Mann K, Rodriguez-Navarro AB, McKee MD (enero de 2012). "La cáscara de huevo: estructura, composición y mineralización". Frontiers in Bioscience . 17 (4): 1266–80. doi : 10.2741/3985 . PMID 22201802.
^ ab McKee MD, Nanci A, Khan SR (diciembre de 1995). "Inmunodetección ultraestructural de osteopontina y osteocalcina como componentes principales de la matriz de los cálculos renales". Journal of Bone and Mineral Research . 10 (12): 1913–29. doi :10.1002/jbmr.5650101211. PMID 8619372. S2CID 37508279.
^ O'Young J, Chirico S, Al Tarhuni N, Grohe B, Karttunen M, Goldberg HA, Hunter GK (2009). "La fosforilación de los péptidos de osteopontina media la adsorción y la incorporación a los cristales de oxalato de calcio". Células, tejidos y órganos . 189 (1–4): 51–55. doi :10.1159/000151724. PMID 18728346. S2CID 5534110.
^ ab Chien YC, Masica DL, Gray JJ, Nguyen S, Vali H, McKee MD (agosto de 2009). "Modulación del crecimiento de oxalato de calcio dihidrato mediante unión selectiva a la cara cristalina de osteopontina fosforilada y péptido de poliaspartato que muestra oclusión por zonificación sectorial (composicional)". The Journal of Biological Chemistry . 284 (35): 23491–501. doi : 10.1074/jbc.M109.021899 . PMC 2749123 . PMID 19581305.
^ Sodek J, Ganss B, McKee MD (2000). "Osteopontina". Revisiones críticas en biología oral y medicina . 11 (3): 279–303. doi : 10.1177/10454411000110030101 . PMID 11021631.
^ ab Reznikov N, Hoac B, Buss DJ, Addison WN, Barros NM, McKee MD (septiembre de 2020). "Estarcido biológico de la mineralización en el esqueleto: eliminación enzimática local de inhibidores en la matriz extracelular". Bone . 138 : 115447. doi :10.1016/j.bone.2020.115447. PMID 32454257. S2CID 218909350.
^ ab McKee MD, Buss DJ, Reznikov N (marzo de 2022). "Teselación mineral en el hueso y el principio de estarcido para la mineralización de la matriz extracelular". Revista de biología estructural . 214 (1): 107823. doi :10.1016/j.jsb.2021.107823. PMID 34915130. S2CID 245187449.
^ McKee MD, Hoac B, Addison WN, Barros NM, Millán JL, Chaussain C (octubre de 2013). "Mineralización de la matriz extracelular en tejidos periodontales: proteínas de la matriz no colágena, enzimas y relación con la hipofosfatasia y la hipofosfatemia ligada al cromosoma X". Periodontología 2000. 63 ( 1): 102–122. doi :10.1111/prd.12029. PMC 3766584. PMID 23931057 .
^ Boukpessi T, Hoac B, Coyac BR, Leger T, Garcia C, Wicart P, et al. (febrero de 2017). "Osteopontina y patobiología dentoósea de la hipofosfatemia ligada al cromosoma X". Bone . 95 : 151–161. doi :10.1016/j.bone.2016.11.019. PMID 27884786.
^ McKee MD, Addison WN, Kaartinen MT (2005). "Jerarquías de la matriz extracelular y la organización mineral en el hueso del complejo craneofacial y el esqueleto". Células, tejidos y órganos . 181 (3–4): 176–188. doi :10.1159/000091379. PMID : 16612083. S2CID : 40705942.
^ ab Steitz SA, Speer MY, McKee MD, Liaw L, Almeida M, Yang H, Giachelli CM (diciembre de 2002). "La osteopontina inhibe la deposición mineral y promueve la regresión de la calcificación ectópica". The American Journal of Pathology . 161 (6): 2035–46. doi :10.1016/S0002-9440(10)64482-3. PMC 1850905 . PMID 12466120.
^ Giachelli CM (marzo de 1999). "Calcificación ectópica: recopilación de datos concretos sobre la mineralización de los tejidos blandos". The American Journal of Pathology . 154 (3): 671–5. doi :10.1016/S0002-9440(10)65313-8. PMC 1866412 . PMID 10079244.
^ Kaartinen MT, Murshed M, Karsenty G, McKee MD (abril de 2007). "Regulación positiva y polimerización de osteopontina por transglutaminasa 2 en arterias calcificadas de ratones deficientes en proteína Matrix Gla". The Journal of Histochemistry and Cytochemistry . 55 (4): 375–386. doi : 10.1369/jhc.6A7087.2006 . PMID 17189522.
^ Choi ST, Kim JH, Kang EJ, Lee SW, Park MC, Park YB, Lee SK (diciembre de 2008). "La osteopontina podría estar involucrada en la remodelación ósea en lugar de en la inflamación en la espondilitis anquilosante". Reumatología . 47 (12): 1775–9. doi : 10.1093/rheumatology/ken385 . PMID 18854347.
^ McKee MD, Nanci A (1996). "Osteopontina: una proteína de la matriz extracelular interfacial en tejidos mineralizados". Connective Tissue Research . 35 (1–4): 197–205. doi :10.3109/03008209609029192. PMID 9084658.
^ Crawford HC, Matrisian LM, Liaw L (noviembre de 1998). "Roles distintos de la osteopontina en la actividad de defensa del huésped y la supervivencia tumoral durante la progresión del carcinoma de células escamosas in vivo". Cancer Research . 58 (22): 5206–15. PMID 9823334.
^ Apte UM, Banerjee A, McRee R, Wellberg E, Ramaiah SK (agosto de 2005). "Función de la osteopontina en la infiltración hepática de neutrófilos durante la esteatohepatitis alcohólica". Toxicología y farmacología aplicada . 207 (1): 25–38. Bibcode :2005ToxAP.207...25A. doi :10.1016/j.taap.2004.12.018. PMID 15885730.
^ Koh A, da Silva AP, Bansal AK, Bansal M, Sun C, Lee H, et al. (diciembre de 2007). "El papel de la osteopontina en la función de los neutrófilos". Inmunología . 122 (4): 466–475. doi :10.1111/j.1365-2567.2007.02682.x. PMC 2266047 . PMID 17680800.
^ Ohshima S, Kobayashi H, Yamaguchi N, Nishioka K, Umeshita-Sasai M, Mima T, et al. (abril de 2002). "Expresión de osteopontina en sitios de erosión ósea en un modelo murino experimental de artritis inducida por colágeno: posible participación de la osteopontina en la destrucción ósea en la artritis". Artritis y reumatismo . 46 (4): 1094–1101. doi : 10.1002/art.10143 . PMID 11953989.
^ Sakata M, Tsuruha JI, Masuko-Hongo K, Nakamura H, Matsui T, Sudo A, et al. (julio de 2001). "Autoanticuerpos contra la osteopontina en pacientes con osteoartritis y artritis reumatoide". The Journal of Rheumatology . 28 (7): 1492–5. PMID 11469452.
^ Burdo TH, Wood MR, Fox HS (junio de 2007). "La osteopontina previene la recirculación de monocitos y la apoptosis". Journal of Leukocyte Biology . 81 (6): 1504–11. doi :10.1189/jlb.1106711. PMC 2490714 . PMID 17369493.
^ Yumoto K, Ishijima M, Rittling SR, Tsuji K, Tsuchiya Y, Kon S, et al. (abril de 2002). "La deficiencia de osteopontina protege las articulaciones contra la destrucción en la artritis inducida por anticuerpos anti-colágeno tipo II en ratones". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 99 (7): 4556–61. Bibcode :2002PNAS...99.4556Y. doi : 10.1073/pnas.052523599 . PMC 123686 . PMID 11930008.
^ Jacobs JP, Pettit AR, Shinohara ML, Jansson M, Cantor H, Gravallese EM, et al. (agosto de 2004). "Falta de requerimiento de osteopontina para la inflamación, erosión ósea y daño del cartílago en el modelo K/BxN de artritis mediada por autoanticuerpos". Arthritis and Rheumatism . 50 (8): 2685–94. doi :10.1002/art.20381. PMID 15334485.
^ Chabas D, Baranzini SE, Mitchell D, Bernard CC, Rittling SR, Denhardt DT, et al. (noviembre de 2001). "La influencia de la citocina proinflamatoria, osteopontina, en la enfermedad desmielinizante autoinmune". Science . 294 (5547): 1731–5. Bibcode :2001Sci...294.1731C. doi :10.1126/science.1062960. PMID 11721059. S2CID 86208485.
^ abcde Xanthou G, Alissafi T, Semitekolou M, Simoes DC, Economidou E, Gaga M, et al. (mayo de 2007). "La osteopontina tiene un papel crucial en la enfermedad alérgica de las vías respiratorias a través de la regulación de los subconjuntos de células dendríticas". Nature Medicine . 13 (5): 570–8. doi :10.1038/nm1580. PMC 3384679 . PMID 17435770.
^ Simoes DC, Xanthou G, Petrochilou K, Panoutsakopoulou V, Roussos C, Gratziou C (mayo de 2009). "La deficiencia de osteopontina protege contra la remodelación de las vías respiratorias y la hiperreactividad en el asma crónico". American Journal of Respiratory and Critical Care Medicine . 179 (10): 894–902. doi :10.1164/rccm.200807-1081OC. PMID 19234104.
^ Samitas K, Zervas E, Vittorakis S, Semitekolou M, Alissafi T, Bossios A, et al. (febrero de 2011). "Expresión de osteopontina y relación con la gravedad de la enfermedad en el asma humano". The European Respiratory Journal . 37 (2): 331–341. doi : 10.1183/09031936.00017810 . PMID 20562127.
^ Hillas G, Loukides S, Kostikas K, Simoes D, Petta V, Konstantellou E, et al. (enero de 2013). "Aumento de los niveles de osteopontina en el sobrenadante del esputo de asmáticos fumadores". Cytokine . 61 (1): 251–5. doi :10.1016/j.cyto.2012.10.002. PMID 23098767.
^ Samitas K, Zervas E, Xanthou G, Panoutsakopoulou V, Gaga M (marzo de 2013). "La osteopontina aumenta en el líquido de lavado broncoalveolar y el tejido bronquial de los asmáticos fumadores". Citocinas . 61 (3): 713–5. doi :10.1016/j.cyto.2012.12.028. PMID 23384656.
^ ab Gassler N, Autschbach F, Gauer S, Bohn J, Sido B, Otto HF, et al. (noviembre de 2002). "Expresión de osteopontina (Eta-1) en la enfermedad de Crohn del íleon terminal". Revista escandinava de gastroenterología . 37 (11): 1286–95. doi :10.1080/003655202761020560. PMID 12465727. S2CID 6381267.
^ Sato T, Nakai T, Tamura N, Okamoto S, Matsuoka K, Sakuraba A, et al. (septiembre de 2005). "La osteopontina/Eta-1 regulada positivamente en la enfermedad de Crohn regula la respuesta inmunitaria Th1". Gut . 54 (9): 1254–62. doi :10.1136/gut.2004.048298. PMC 1774642 . PMID 16099792.
^ Mishima R, Takeshima F, Sawai T, Ohba K, Ohnita K, Isomoto H, et al. (febrero de 2007). "Niveles elevados de osteopontina plasmática en pacientes con enfermedad inflamatoria intestinal". Journal of Clinical Gastroenterology . 41 (2): 167–172. doi :10.1097/MCG.0b013e31802d6268. PMID 17245215. S2CID 25039321.
^ ab Kourepini E, Aggelakopoulou M, Alissafi T, Paschalidis N, Simoes DC, Panoutsakopoulou V (marzo de 2014). "La expresión de osteopontina por las células dendríticas CD103- impulsa la inflamación intestinal". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 111 (9): E856–E865. Bibcode :2014PNAS..111E.856K. doi : 10.1073/pnas.1316447111 . PMC 3948306 . PMID 24550510.
^ Steinman L (febrero de 2007). "Una breve historia de T(H)17, la primera revisión importante de la hipótesis T(H)1/T(H)2 del daño tisular mediado por células T". Nature Medicine . 13 (2): 139–145. doi :10.1038/nm1551. PMID 17290272. S2CID 10640379.
^ Clay R, Siddiqi SA (enero de 2019). "Avances recientes en diagnóstico molecular y dianas terapéuticas para el cáncer de páncreas". Enfoque teranóstico para el cáncer de páncreas : 325–367. doi :10.1016/B978-0-12-819457-7.00016-5. ISBN978-0-12-819457-7.S2CID214323538 .
^ Clay R, Siddiqi SA (1 de enero de 2019). "Avances recientes en diagnóstico molecular y dianas terapéuticas para el cáncer de páncreas". Enfoque teranóstico para el cáncer de páncreas : 325–367. doi :10.1016/B978-0-12-819457-7.00016-5. ISBN978-0-12-819457-7.S2CID214323538 .
^ Mori R, Shaw TJ, Martin P (enero de 2008). "Mecanismos moleculares que vinculan la inflamación de las heridas y la fibrosis: la reducción de la osteopontina conduce a una reparación rápida y a una reducción de la cicatrización". The Journal of Experimental Medicine . 205 (1): 43–51. doi :10.1084/jem.20071412. PMC 2234383 . PMID 18180311.
"Gel para acelerar la cicatrización de heridas". BBC News . 22 de enero de 2008.
^ Shojaei F, Scott N, Kang X, Lappin PB, Fitzgerald AA, Karlicek S, et al. (marzo de 2012). "La osteopontina induce el crecimiento de tumores metastásicos en un modelo preclínico de cáncer de pulmón no microcítico". Journal of Experimental & Clinical Cancer Research . 31 (1): 26. doi : 10.1186/1756-9966-31-26 . PMC 3325875 . PMID 22444159.
^ Farrokhi V, Chabot JR, Neubert H, Yang Z (mayo de 2018). "Evaluación de la viabilidad de neutralizar la osteopontina con diversas modalidades de anticuerpos terapéuticos". Scientific Reports . 8 (1): 7781. Bibcode :2018NatSR...8.7781F. doi :10.1038/s41598-018-26187-w. PMC 5958109 . PMID 29773891.
^ Leavenworth JW, Verbinnen B, Wang Q, Shen E, Cantor H (enero de 2015). "La osteopontina intracelular regula la homeostasis y la función de las células asesinas naturales". Proc Natl Acad Sci USA . 112 (2): 494–9. Bibcode :2015PNAS..112..494L. doi : 10.1073/pnas.1423011112 . PMC 4299239 . PMID 25550515.
^ Drobits B, Holcmann M, Amberg N, Swiecki M, Grundtner R, Hammer M, Colonna M, Sibilia M (febrero de 2012). "Imiquimod elimina tumores en ratones independientemente de la inmunidad adaptativa al convertir las pDC en células efectoras que destruyen tumores". J Clin Invest . 122 (2): 575–85. doi :10.1172/JCI61034. PMC 3266798 . PMID 22251703.
^ Simoes DC, Paschalidis N, Kourepini E, Panoutsakopoulou V (septiembre de 2022). "Un eje de integrina induce la producción de IFN-β en células dendríticas plasmocitoides". J Cell Biol . 221 (9): e202102055. doi :10.1083/jcb.202102055. PMC 9354318 . PMID 35878016.
^ Singh M, Dalal S, Singh K (noviembre de 2014). "Osteopontina: en la encrucijada de la supervivencia de los miocitos y la función miocárdica". Ciencias de la vida . 118 (1): 1–6. doi :10.1016/j.lfs.2014.09.014. PMC 4254317 . PMID 25265596.
^ ab Singh M, Foster CR, Dalal S, Singh K (marzo de 2010). "Osteopontina: papel en la deposición de la matriz extracelular y la remodelación miocárdica después de un infarto de miocardio". Journal of Molecular and Cellular Cardiology . 48 (3): 538–543. doi :10.1016/j.yjmcc.2009.06.015. PMC 2823840 . PMID 19573532.
^ Shirakawa K, Sano M (julio de 2021). "Osteopontina en enfermedades cardiovasculares". Biomolecules . 11 (7): 1047. doi : 10.3390/biom11071047 . PMC 8301767 . PMID 34356671.
^ Graf K, Do YS, Ashizawa N, Meehan WP, Giachelli CM, Marboe CC, et al. (noviembre de 1997). "La expresión de osteopontina miocárdica está asociada con la hipertrofia ventricular izquierda". Circulation . 96 (9): 3063–71. doi :10.1161/01.CIR.96.9.3063. PMID 9386176.
^ Maetzler W, Berg D, Schalamberidze N, Melms A, Schott K, Mueller JC, et al. (marzo de 2007). "La osteopontina está elevada en la enfermedad de Parkinson y su ausencia conduce a una neurodegeneración reducida en el modelo MPTP". Neurobiología de la enfermedad . 25 (3): 473–482. doi :10.1016/j.nbd.2006.10.020. PMID 17188882. S2CID 30275400.
^ Porter JD, Khanna S, Kaminski HJ, Rao JS, Merriam AP, Richmonds CR, et al. (febrero de 2002). "Una respuesta inflamatoria crónica domina la firma molecular del músculo esquelético en ratones mdx deficientes en distrofina". Human Molecular Genetics . 11 (3): 263–272. doi : 10.1093/hmg/11.3.263 . PMID 11823445.
^ Haslett JN, Sanoudou D, Kho AT, Bennett RR, Greenberg SA, Kohane IS, et al. (noviembre de 2002). "Comparación de la expresión génica en biopsias de distrofia muscular de Duchenne (DMD) y músculo esquelético normal". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 99 (23): 15000–5. Bibcode :2002PNAS...9915000H. doi : 10.1073/pnas.192571199 . PMC 137534 . PMID 12415109.
^ Hirata A, Masuda S, Tamura T, Kai K, Ojima K, Fukase A, et al. (julio de 2003). "Perfiles de expresión de citocinas y genes relacionados en la regeneración del músculo esquelético después de la inyección de cardiotoxina: un papel para la osteopontina". The American Journal of Pathology . 163 (1): 203–215. doi :10.1016/S0002-9440(10)63644-9. PMC 1868192 . PMID 12819025.
^ Vetrone SA, Montecino-Rodriguez E, Kudryashova E, Kramerova I, Hoffman EP, Liu SD, et al. (junio de 2009). "La osteopontina promueve la fibrosis en el músculo distrófico del ratón modulando los subconjuntos de células inmunes y el TGF-beta intramuscular". The Journal of Clinical Investigation . 119 (6): 1583–94. doi :10.1172/JCI37662. PMC 2689112 . PMID 19451692.
^ Pegoraro E, Hoffman EP, Piva L, Gavassini BF, Cagnin S, Ermani M, et al. (Enero de 2011). "El genotipo SPP1 es un determinante de la gravedad de la enfermedad en la distrofia muscular de Duchenne". Neurología . 76 (3): 219–226. doi :10.1212/WNL.0b013e318207afeb. PMC 3034396 . PMID 21178099.
^ El Deeb S, Abdelnaby R, Khachab A, Bläsius K, Tingart M, Rath B (julio de 2016). "La osteopontina como marcador bioquímico e indicador de gravedad de la osteoartritis idiopática de la cadera". Hip International . 26 (4): 397–403. doi :10.5301/hipint.5000361. PMID 27229171. S2CID 40819265.
^ Kang YJ, Forbes K, Carver J, Aplin JD (abril de 2014). "El papel de la interacción osteopontina-integrina αvβ3 en la implantación: análisis funcional utilizando tres modelos in vitro diferentes". Human Reproduction . 29 (4): 739–749. doi :10.1093/humrep/det433. PMID 24442579.
^ Johnson GA, Burghardt RC, Bazer FW, Spencer TE (noviembre de 2003). "Osteopontina: funciones en la implantación y placentación". Biología de la reproducción . 69 (5): 1458–71. doi : 10.1095/biolreprod.103.020651 . PMID 12890718.
Lectura adicional
Fujisawa R (marzo de 2002). "[Avances recientes en la investigación sobre proteínas de la matriz ósea]". Nihon Rinsho. Revista japonesa de medicina clínica . 60. 60 (Supl. 3): 72–78. PMID 11979972.
Denhardt DT, Mistretta D, Chambers AF, Krishna S, Porter JF, Raghuram S, Rittling SR (2003). "Regulación transcripcional de la osteopontina y el fenotipo metastásico: evidencia de un potenciador activado por Ras en el promotor humano de OPN". Clinical & Experimental Metastasis . 20 (1): 77–84. doi :10.1023/A:1022550721404. PMID 12650610. S2CID 20286402.
Yeatman TJ, Chambers AF (2003). "Osteopontina y progresión del cáncer de colon". Clinical & Experimental Metastasis . 20 (1): 85–90. doi :10.1023/A:1022502805474. PMID 12650611. S2CID 25253392.
O'Regan A (diciembre de 2003). "El papel de la osteopontina en la enfermedad pulmonar". Cytokine & Growth Factor Reviews . 14 (6): 479–488. doi :10.1016/S1359-6101(03)00055-8. PMID 14563350.
Wai PY, Kuo PC (octubre de 2004). "El papel de la osteopontina en la metástasis tumoral". The Journal of Surgical Research . 121 (2): 228–241. doi :10.1016/j.jss.2004.03.028. PMID 15501463.
Konno S, Hizawa N, Nishimura M, Huang SK (diciembre de 2006). "Osteopontina: un biomarcador potencial para la inmunoterapia exitosa con veneno de abeja y una molécula potencial para inhibir las respuestas alérgicas mediadas por IgE". Allergology International . 55 (4): 355–9. doi : 10.2332/allergolint.55.355 . PMID 17130676.
Rodrigues LR, Teixeira JA, Schmitt FL, Paulsson M, Lindmark-Mänsson H (junio de 2007). "El papel de la osteopontina en la progresión tumoral y la metástasis en el cáncer de mama". Epidemiología del cáncer, biomarcadores y prevención . 16 (6): 1087–97. doi : 10.1158/1055-9965.EPI-06-1008 . hdl : 1822/7274 . PMID: 17548669.
Ramaiah SK, Rittling S (agosto de 2007). "El papel de la osteopontina en la regulación de las respuestas inflamatorias hepáticas y la lesión hepática tóxica". Opinión de expertos sobre el metabolismo y la toxicología de fármacos . 3 (4): 519–526. doi :10.1517/17425225.3.4.519. PMID 17696803.