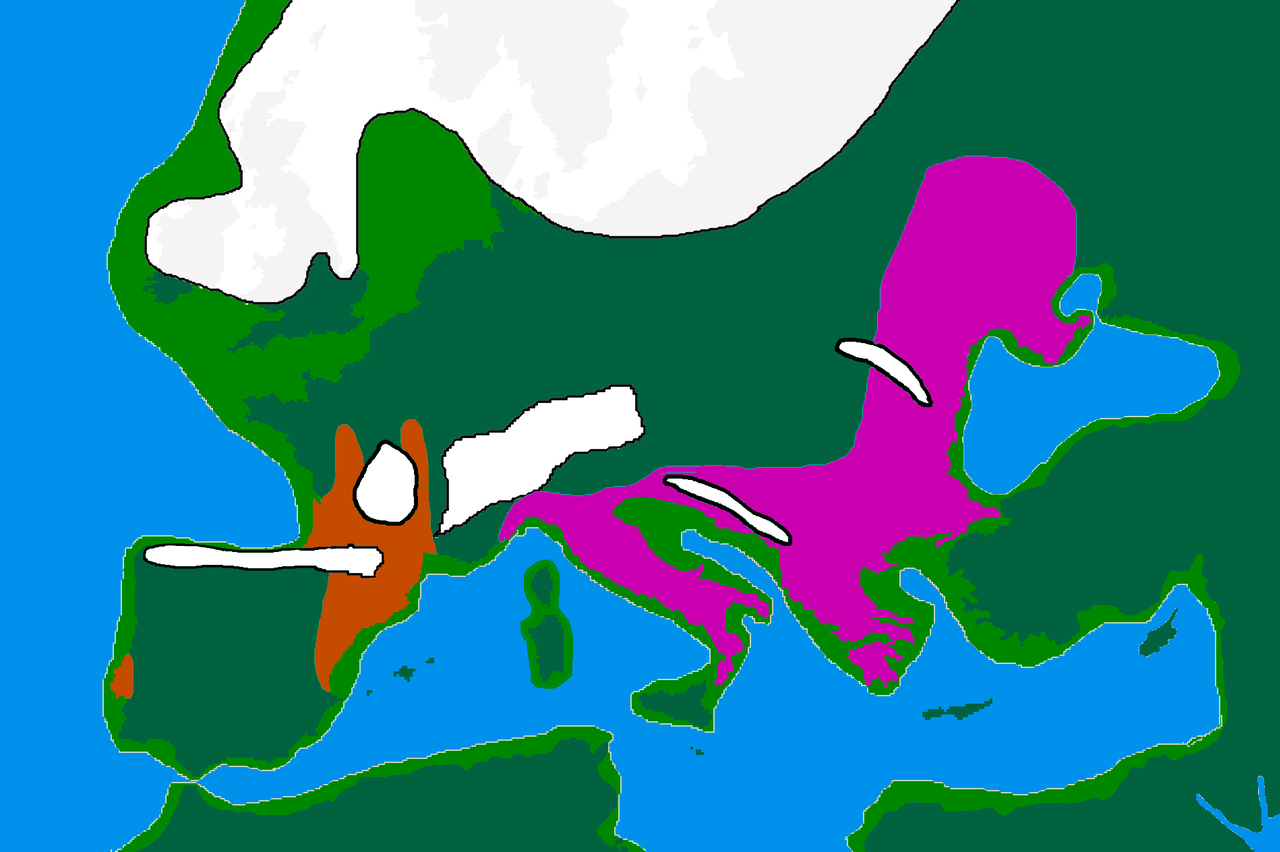
El haplogrupo I probablemente surgió en Europa, [1] [2] y hasta ahora se ha encontrado en yacimientos paleolíticos en toda Europa, pero no fuera de ella. [3] Se separó de su ancestro común IJ* hace unos 43.000 años. [4] Se ha encontrado evidencia temprana del haplogrupo J en el Cáucaso e Irán. [3] Además, se han encontrado ejemplos vivos del precursor Haplogrupo IJ* solo en Irán, entre los Mazandarani y los persas étnicos de Fars . [5] Esto puede indicar que IJ se originó en el suroeste de Asia .
El ejemplo más antiguo encontrado fue originalmente el de Paglicci133 de Italia , que tiene al menos 31.000 años de antigüedad, [6] sin embargo, en un estudio posterior esto se cambió, y en su lugar se informó que Dolní Věstonice (DV14) de la República Checa era el más antiguo, con al menos 30.800 años de antigüedad. [7]
El haplogrupo I se ha encontrado en múltiples individuos pertenecientes a la cultura gravetiense. Los gravetienses se expandieron hacia el oeste desde el extremo más alejado de Europa del Este, probablemente Rusia, hasta Europa Central. Están asociados con un grupo genético que normalmente se denomina grupo de Věstonice. [8] [9] [10]
La evidencia disponible sugiere que I-M170 fue precedido en áreas en las que luego se volvería dominante por los haplogrupos K2a (K-M2308) y C1 (Haplogrupo C-F3393). K2a y C1 se han encontrado en los restos masculinos secuenciados más antiguos de Eurasia occidental (que datan de alrededor de 45.000 a 35.000 años BP ), como: hombre de Ust'-Ishim (moderna Siberia occidental) K2a*, Oase 1 (Rumania) K2a*, Kostenki 14 (suroeste de Rusia) C1b, y Goyet Q116-1 (Bélgica) C1a. [3] [11] El I-M170 más antiguo encontrado es el de un individuo conocido como Krems WA3 (Baja Austria), que data de alrededor de 33.000-24.000 BP. En el mismo sitio, también se encontraron dos niños gemelos, ambos fueron asignados al haplogrupo I*. [12] [13]
El haplogrupo IJ estaba en Oriente Medio y/o Europa hace unos 40.000 años. [ cita requerida ] El TMRCA (tiempo hasta el ancestro común más reciente ) para I-M170 fue estimado por Karafet y colegas en 2008 en hace 22.200 años, con un intervalo de confianza entre 15.300 y 30.000 años atrás. [4] Esto haría que el evento fundador de I-M170 fuera aproximadamente contemporáneo con el Último Máximo Glacial (LGM), que duró desde hace 26.500 años hasta hace aproximadamente 19.500 años. [14] TMRCA es una estimación del tiempo de divergencia del subclado. Rootsi y colegas en 2004 también señalan otras dos fechas para un clado, la edad de variación de STR y el tiempo transcurrido desde la divergencia de la población. Estas dos últimas fechas están aproximadamente asociadas y ocurren algo después de la divergencia del subclado. Para el haplogrupo I-M170, estiman que el tiempo hasta la variación de STR fue hace 24.000 ± 7.100 años y el tiempo hasta la divergencia de la población fue hace 23.000 ± 7.700 años. [1] Estas estimaciones son consistentes con las de Karafet 2008 citadas anteriormente. Sin embargo, Underhill y sus colegas calculan que el tiempo hasta la divergencia de los subclados I1 e I2 fue hace 28.400 ± 5.100 años, aunque calculan la edad de variación de STR de I1 en solo 8.100 ± 1.500 años. [15]
Semino (2000) especuló que la dispersión inicial de esta población corresponde a la difusión de la cultura Gravetiense . [16] Posteriormente el haplogrupo, junto con dos casos del Haplogrupo C , fue encontrado en restos humanos pertenecientes a la cultura Gravetiense mencionada anteriormente y en individuos de las culturas Magdaleniense y Aziliense . [17] Rootsi y colegas en 2004 sugirieron que cada una de las poblaciones ancestrales ahora dominadas por un subclado particular del Haplogrupo I-M170 experimentó una expansión poblacional independiente inmediatamente después del Último Máximo Glacial. [1]
Los cinco casos conocidos del haplogrupo I en restos humanos europeos del Paleolítico superior lo convierten en uno de los haplogrupos más frecuentes de ese período. [17] En 2016, se descubrió que los restos de un cazador-recolector de 31.210 a 34.580 años de antigüedad de la cueva de Paglicci , Apulia, Italia, contenían I-M170. [18] Hasta ahora, solo se han documentado el haplogrupo F* y el haplogrupo C 1b, una vez cada uno, en restos más antiguos en Europa. El subclade I2 de I-M170 es el haplogrupo principal encontrado en restos masculinos en la Europa mesolítica , hasta alrededor del 6000 a. C., cuando se produjo una migración masiva a Europa de agricultores anatolios que portaban el ADN-Y G2a. [19]
La documentación más antigua de I1 procede del Neolítico húngaro, aunque debe haberse separado de I2 en un momento anterior.
En un caso, el haplogrupo I fue encontrado lejos de Europa, entre restos de Mongolia de 2.000 años de antigüedad. [20]
Parecería ser que olas separadas de movimiento de población impactaron el sudeste de Europa . El papel de los Balcanes como un corredor de larga data hacia Europa desde Anatolia y/o el Cáucaso se demuestra por los orígenes filogenéticos comunes de ambos haplogrupos I y J en el haplogrupo parental IJ (M429). Esta ascendencia común sugiere que los subclados de IJ ingresaron a los Balcanes desde Anatolia o el Cáucaso, algún tiempo antes del Último Máximo Glacial. I y J se distribuyeron posteriormente en Asia y Europa en un patrón filogeográfico disyuntivo típico de los haplogrupos "hermanos". Es probable que un corredor geográfico natural como los Balcanes haya sido utilizado más tarde por miembros de otros subclados de IJ, así como otros haplogrupos, incluidos los asociados con los primeros agricultores europeos.
La existencia del haplogrupo IJK (antecesor de los haplogrupos IJ y K (M9)) y su distancia evolutiva con respecto a otros subclados del haplogrupo F (M89) respaldan la inferencia de que los haplogrupos IJ y K surgieron en el sudoeste de Asia. Se han registrado portadores vivos de F* e IJ* en la meseta iraní . [5]
Distribución
Frecuencias del haplogrupo I:
Subgrupos
Los subclados del haplogrupo I-M170 con sus mutaciones definitorias, a partir de 2011. [86] Los árboles filogenéticos actualizados que enumeran todos los subclados conocidos actualmente de I se pueden encontrar en Y-Full y FamilyTreeDNA.
I-M170 (L41, M170, M258, P19_1, P19_2, P19_3, P19_4, P19_5, P38, P212, Página123, U179) Oriente Medio, Cáucaso, Europa.
Haplogrupo I1 de I-M253 (L64, L75, L80, L81, L118, L121/S62, L123, L124/S64, L125/S65, L157, L186, L187, M253, M307.2/P203.2, M450/S109, P30, P40, S63, S66, S107, S108, S110, S111) Típico de poblaciones de Escandinavia y el noroeste de Europa , con una distribución moderada en toda Europa del Este en Anatolia en un 1% [87]
Tenga en cuenta que el nombre de algunos de los subgrupos ha cambiado a medida que se han identificado nuevos marcadores y la secuencia de mutaciones se ha vuelto más clara.
Yo-M170
El subclado compuesto I-M170 contiene individuos que descienden directamente de los primeros miembros del Haplogrupo I y no presentan ninguna de las mutaciones posteriores que identifican a los subclados nombrados restantes.
Se han encontrado varios individuos I*, que no pertenecen a ningún subclado conocido, entre el pueblo Lak de Daguestán , a una tasa de (3/21), [89] así como en Turquía (8/741), Adiguesia en el Cáucaso (2/138) e Irak (1/176), aunque I-M170 se presenta solo en frecuencias muy bajas entre las poblaciones modernas de estas regiones en su conjunto. Esto es consistente con la creencia de que el haplogrupo apareció por primera vez en el suroeste de Eurasia.
(Ninguno de los estudios de los que se extrajeron las cifras anteriores excluyó el clado I2-M438 actual en su totalidad, sino sólo ciertos subclados, por lo que estos presuntos casos I* posiblemente pertenezcan a I2).
También se ha descubierto que un macho hazara vivo de Afganistán es portador de I*, y se han descartado todos los subclados conocidos de I1 (M253) e I2 (M438). [91]
I1-M253
El haplogrupo I1-M253 (M253, M307, P30, P40) muestra un gradiente de frecuencia muy claro, con una frecuencia máxima de aproximadamente el 35% entre las poblaciones del sur de Noruega, el suroeste de Suecia y Dinamarca, y frecuencias que disminuyen rápidamente hacia los bordes del mundo históricamente influenciado por los germánicos . Una excepción notable es Finlandia, donde la frecuencia en los finlandeses occidentales es de hasta el 40%, y en ciertas provincias como Satakunta más del 50%. Se cree que I1 se volvió común como resultado de un efecto fundador durante la Edad del Bronce Nórdica , y posteriormente se extendió por toda Europa durante el Período de Migración cuando las tribus germánicas migraron desde el sur de Escandinavia y el norte de Alemania a otros lugares de Europa. [92]
Fuera de Fennoscandia , la distribución del haplogrupo I1-M253 está estrechamente correlacionada con la del haplogrupo I2a2-M436; pero entre los escandinavos (incluidos los pueblos germánicos y urálicos de la región) casi todos los cromosomas Y del haplogrupo I-M170 son I1-M253. Otra característica de los cromosomas Y escandinavos I1-M253 es su diversidad de haplotipos bastante baja (diversidad STR): se ha encontrado una mayor variedad de cromosomas Y del haplogrupo I1-M253 entre los franceses e italianos , a pesar de la frecuencia general mucho menor del haplogrupo I1-M253 entre las poblaciones francesa e italiana modernas. Esto, junto con la estructura del árbol filogenético de I1-M253, sugiere firmemente que la mayoría de los varones I1 vivos son descendientes de un grupo inicialmente pequeño de hombres reproductivamente exitosos que vivieron en Escandinavia durante la Edad del Bronce Nórdica. [93] [94]
I2-M438
El haplogrupo I2-M438 , anteriormente I1b , puede haberse originado en el sur de Europa (ahora se encuentra en sus frecuencias más altas en los Balcanes occidentales y Cerdeña ) hace unos 15.000–17.000 años y se desarrolló en tres subgrupos principales: I2-M438*, I2a-L460, I2b-L415 e I2c-L596.
I2a1a-M26
El haplogrupo I2a1a-M26 se destaca por su fuerte presencia en Cerdeña. El haplogrupo I-M170 comprende aproximadamente el 40% de todas las líneas paternas entre los sardos , y el I2a1a-M26 es el tipo predominante de I entre ellos.
El haplogrupo I2a1a-M26 está prácticamente ausente al este de Francia e Italia, [95] mientras que se encuentra en frecuencias bajas pero significativas fuera de Cerdeña en las Islas Baleares , Castilla y León , el País Vasco , los Pirineos , el sur y oeste de Francia y partes del Magreb en el norte de África, Gran Bretaña e Irlanda. El haplogrupo I2a1a-M26 parece ser el único subclado del haplogrupo I-M170 encontrado entre los vascos , pero parece encontrarse en frecuencias algo más altas entre las poblaciones generales de Castilla y León en España y Béarn en Francia que entre la población de vascos étnicos. [ cita requerida ] La mutación M26 se encuentra en varones nativos que habitan en todas las regiones geográficas donde se pueden encontrar megalitos, incluidas regiones tan lejanas y culturalmente desconectadas como las Islas Canarias, las Islas Baleares, Córcega, Irlanda y Suecia. [95]
Distribución aproximada de frecuencia y varianza de los grupos del haplogrupo I-P37, ancestral "Dnieper-Carpathian" (DYS448=20) y derivado "Balkan" (DYS448=19: representado por un único SNP I-PH908), en Europa del Este según OM Utevska (2017).
El haplogrupo I2a1b-M423 es el haplogrupo I-M170 del cromosoma Y más frecuente en las poblaciones de Europa central y oriental, alcanzando su pico en los Balcanes occidentales , sobre todo en Dalmacia (50-60% [33] ) y Bosnia-Herzegovina (hasta 71%, [96] promedio 40-50% [33] ). Su subclade I-L161 tiene mayor varianza en Irlanda y Gran Bretaña, pero la frecuencia general es muy baja (2-3%), mientras que el subclade I-L162 tiene la varianza más alta y también una alta concentración en Europa del Este (Ucrania, sureste de Polonia, Bielorrusia). [97]
I2a2-M436
La distribución del haplogrupo I2a2-M436 (M436/P214/S33, P216/S30, P217/S23, P218/S32) está estrechamente relacionada con la del haplogrupo I1, excepto en Fennoscandia , lo que sugiere que probablemente lo albergaba al menos una de las poblaciones de refugio del Paleolítico que también albergaba el haplogrupo I1-M253; la falta de correlación entre las distribuciones de I1-M253 e I2a2-M436 en Fennoscandia puede ser el resultado de que el haplogrupo I2a2-M436 se vio más fuertemente afectado en el asentamiento más temprano de esta región por los efectos fundadores y la deriva genética debido a su rareza, ya que el haplogrupo I2a2-M436 comprende menos del 10% de la diversidad total del cromosoma Y de todas las poblaciones fuera de Baja Sajonia . El haplogrupo I2a2-M436 se ha encontrado en más del 4% de la población solo en Alemania, los Países Bajos, Bélgica, Dinamarca, Inglaterra (sin incluir Cornualles ), Escocia y los extremos meridionales de Suecia y Noruega en el noroeste de Europa; las provincias de Normandía , Maine , Anjou y Perche en el noroeste de Francia; la provincia de Provenza en el sureste de Francia; las regiones de Toscana , Umbría y Lacio en Italia; y Moldavia y el área alrededor del óblast de Riazán y la República de Mordovia en Rusia en Europa del Este. Un subclado del haplogrupo I2a2-M436, concretamente I2a2a1a1-M284, se ha encontrado casi exclusivamente entre la población de Gran Bretaña, lo que se ha tomado como sugerencia de que el clado puede tener una historia muy larga en esa isla. Es notable, sin embargo, que las distribuciones del haplogrupo I1-M253 y el haplogrupo I2a2-M436 parecen correlacionarse bastante bien con el alcance de la influencia histórica de los pueblos germánicos . La presencia puntual de ambos haplogrupos con una baja frecuencia en el área de las regiones históricas de Bitinia y Galacia en Turquía puede estar relacionada con la Guardia Varega o más bien sugiere una conexión con los antiguos galos de Tracia , varias tribus de las cuales se registra que inmigraron a esas partes de Anatolia por invitación de Nicomedes I de Bitinia . Esta sugerencia está respaldada por estudios genéticos recientes sobre el haplogrupo I2b2-L38 de ADN-Y que han concluido que hubo alguna migración de la Edad del Hierro tardía del pueblo celta La Tène, a través de Bélgica, a las Islas Británicas, incluido el noreste de Irlanda. [98]
El haplogrupo I2a2-M436 también se presenta en aproximadamente el 1% de los sardos y en los hazaras de Afganistán en un 3%. [99]
Especificaciones de la mutación
Los detalles técnicos del U179 son:
Cambio de nucleótido (rs2319818): G a A
Posición (par de bases): 275
Tamaño total (pares de bases): 220
Adelante 5′→ 3′:aaggggatatgacgactgatt
Retroceso 5′→ 3′:cagctcctcttttcaactctca
Altura
Este haplogrupo alcanza su frecuencia máxima en los Balcanes occidentales (con la mayor concentración de I2 en la actual Herzegovina ). Puede estar asociado con varones inusualmente altos, ya que se ha informado que los de los Alpes Dináricos son los más altos del mundo, con una altura masculina promedio del rango de 180 cm (5 pies 11 pulgadas) - 182 cm (6 pies 0 pulgadas) en los cantones de Bosnia , 184 cm (6 pies 0 pulgadas) en Sarajevo , 182 cm (6 pies 0 pulgadas) - 186 cm (6 pies 1 pulgada) en los cantones de Herzegovina poblados principalmente por croatas . [100] Un estudio de 2014 que examinó la correlación entre los haplogrupos de ADN-Y y la altura encontró una correlación entre los haplogrupos I1 , R1b-U106, I2a1b-M423 y varones altos. [101] El estudio incluyó las estaturas medias medidas de hombres jóvenes alemanes, suecos, holandeses, daneses, serbios y bosnios. La estatura media de los hombres alemanes fue de 180,2 cm, la de los hombres suecos de 181,4 cm, la de los hombres holandeses de 183,8 cm, la de los hombres daneses de 180,6 cm, la de los serbios de 180,9 cm y la de los hombres croatas de Bosnia de Herzegovina de 185,2 centímetros.
Barać L, Pericić M, Klarić IM, et al. (julio de 2003). "Herencia cromosómica Y de la población croata y sus aislamientos insulares" (PDF) . Eur. J. Hum. Genet . 11 (7): 535–42. doi : 10.1038/sj.ejhg.5200992 . PMID 12825075. S2CID 15822710. Archivado desde el original (PDF) el 2012-12-17 . Consultado el 2005-12-08 .
Bennett, EA, Prat, S., Péan, S., Crépin, L., Yanevich, A., Puaud, S., ... y Geigl, EM (2019). El origen de los Gravetienses: evidencia genómica de un europeo oriental de 36.000 años de antigüedad. BioRxiv, 685404.
Rootsi, S, Kivisild, T, Benuzzi, G, Help, H, et al. (2019). "La filogeografía del haplogrupo I del cromosoma Y revela dominios distintos del flujo genético prehistórico en Europa". Am. J. Hum. Genet . 75 (1): 128–137. doi : 10.1086/422196 . hdl : 10400.13/3045 . PMID: 12825075. S2CID : 2834639.
El Proyecto Genográfico, National Geographic, Atlas del viaje humano
ISOGG, Haplogrupo I del ADN-Y y sus subclados
Referencias
^ a b C Rootsi Siiri; Kivisild, Toomas; Benuzzi, Giorgia; Ayuda, Hela; Bermisheva, Marina; Kutuev, Ildus; Barać, Lovorka; Pericic, Marijana; Balanovsky, Oleg; Pshenichnov, Andrey; Dión, Daniel; Grobei, Mónica; Zhivotovsky, Lev A.; Battaglia, Vincenza; Aquiles, Alejandro; Al-Zahery, Nadia; Parik, Juri; Rey, Roy; Cinnioğlu, Cengiz; Khusnutdinova, Elsa; Rudán, Pavao; Balanovska, Elena; Scheffrahn, Wolfgang; Simonescu, Maya; Brehm, Antonio; Gonçalves, Rita; Rosa, Alejandra; Moisan, Jean-Paul; Chaventre, André; Ferak, Vladimir; et al. (2004). "La filogeografía del haplogrupo I-M170 del cromosoma Y revela dominios distintos del flujo genético prehistórico en Europa". American Journal of Human Genetics . 75 (1): 128–137. doi :10.1086/422196. PMC 1181996 . PMID 15162323.
^ Львович, Рожанский Игорь (2021). "ОБЗОР ДАННЫХ ИСКОПАЕМОЙ ДНК: ГАПЛОКАРТЫ G И I". Formato histórico (4 (28)): 125–140.
^ abc Fu, Qiaomei; et al. (2016). "La historia genética de la Edad de Hielo en Europa". Nature . 534 (7606): 200–5. Bibcode :2016Natur.534..200F. doi :10.1038/nature17993. PMC 4943878 . PMID 27135931.
^ ab Karafet TM, Mendez FL, Meilerman MB, Underhill PA, Zegura SL, Hammer MF (2008). "Nuevos polimorfismos binarios remodelan y aumentan la resolución del árbol del haplogrupo del cromosoma Y humano". Genome Research . 18 (5): 830–8. doi :10.1101/gr.7172008. PMC 2336805 . PMID 18385274.
^ ab Grugni (2012). "Antiguos acontecimientos migratorios en Oriente Medio: nuevas pistas a partir de la variación del cromosoma Y de los iraníes modernos". PLOS ONE . 7 (7): e41252. Bibcode :2012PLoSO...741252G. doi : 10.1371/journal.pone.0041252 . PMC 3399854 . PMID 22815981.
^ Fu, Qiaomei; et al. (2016). "La historia genética de la Edad de Hielo en Europa". Nature . 534 (7606): 200–5. Bibcode :2016Natur.534..200F. doi :10.1038/nature17993. PMC 4943878 . PMID 27135931.
^ Posth, Cosme; Yu, él; Ghalichi, Ayshin; Rougier, Hélène; Crévecoeur, Isabelle; Huang, Yilei; Ringbauer, Harald; Rohrlach, Adam B.; Nägele, Kathrin; Villalba-Mouco, Vanessa; Radzeviciute, Rita; Ferraz, Tiago; Stoessel, Alejandro; Tujbatova, Rezeda; Drucker, Dorothée G. (1 de marzo de 2023). "Paleogenómica del Paleolítico superior al Neolítico cazadores-recolectores europeos". Naturaleza . 615 (7950): 117–126. doi :10.1038/s41586-023-05726-0. hdl : 10256/23099 . ISSN 1476-4687.
^ "Vista detallada de publicaciones". fgga.univie.ac.at . Consultado el 15 de diciembre de 2020 .
^ Mounier, Aurélien; Heuzé, Yann; Samsel, Mathilde; Vasilyev, Sergey; Klaric, Laurent; Villotte, Sébastien (14 de diciembre de 2020). "Morfología craneal gravetiense y afinidades grupales humanas durante el Paleolítico superior europeo". Scientific Reports . 10 (1): 21931. Bibcode :2020NatSR..1021931M. doi : 10.1038/s41598-020-78841-x . ISSN 2045-2322. PMC 7736346 . PMID 33318530.
^ Bennett, E. Andrés; Prat, Sandrine; Péan, Stéphane; Crépin, Laurent; Yanevich, Alexandr; Puaud, Simón; Grange, Thierry; Geigl, Eva-Maria (2 de julio de 2019). "El origen de los gravetienses: evidencia genómica de un europeo del este de 36.000 años". bioRxiv : 685404. doi : 10.1101/685404. S2CID 198249005.
^ Seguin-Orlando et al. (2014) 「Estructura genómica en europeos que data de al menos 36.200 años atrás」
^ Teschler-Nicola, María; Fernández, Daniel; Händel, Marc; Einwögerer, Thomas; Simón, Ulrich; Neugebauer-Maresch, Christine; Tangl, Stefan; Heimel, Patricio; Dobsak, Toni; Retzmann, Anika; Prohaska, Thomas (6 de noviembre de 2020). "El ADN antiguo revela gemelos recién nacidos monocigóticos del Paleolítico superior". Biología de las Comunicaciones . 3 (1): 650.doi : 10.1038 /s42003-020-01372-8 . ISSN 2399-3642. PMC 7648643 . PMID 33159107.
^ "El Y-SNP pide el Krems WA3". Genetiker . 2016-05-11 . Consultado el 2020-12-15 .
^ Clark PU, Dyke AS, Shakun JD, et al. (agosto de 2009). "El último máximo glacial". Science . 325 (5941): 710–4. Bibcode :2009Sci...325..710C. doi :10.1126/science.1172873. PMID 19661421. S2CID 1324559.
^ PA Underhill, NM Myres, S. Rootsi, CT Chow, AA Lin, RP Otillar, R. King, LA Zhivotovsky, O. Balanovsky, A. Pshenichnov, KH Ritchie, LL Cavalli-Sforza, T. Kivisild, R. Villems, SR Woodward, Nuevas relaciones filogenéticas para el haplogrupo I del cromosoma Y: reevaluación de su filogeografía y prehistoria, en P. Mellars, K. Boyle, O. Bar-Yosef y C. Stringer (eds.), Rethinking the Human Evolution (2007), págs. 33-42.
^ Semino O, Passarino G, Oefner PJ, et al. (noviembre de 2000). "El legado genético del Homo sapiens sapiens paleolítico en los europeos actuales: una perspectiva del cromosoma Y". Science . 290 (5494): 1155–9. Bibcode :2000Sci...290.1155S. doi :10.1126/science.290.5494.1155. PMID 11073453.
^ ab "ADN paleolítico de Eurasia". Archivado desde el original el 3 de octubre de 2016. Consultado el 5 de octubre de 2016 .
^ Fu, Qiaomei; Posth, Cosimo; Hajdinjak, Mateja; Petr, Martín; Mallick, Swapan; Fernández, Daniel; Furtwängler, Anja; Haak, Wolfgang; Meyer, Matías (2016). "La historia genética de la Europa de la Edad del Hielo". Naturaleza . 534 (7606): 200–205. Código Bib :2016Natur.534..200F. doi : 10.1038/naturaleza17993. hdl :10211.3/198594. ISSN 0028-0836. PMC 4943878 . PMID 27135931.
^ "El ADN antiguo revela una 'continuidad genética' entre las poblaciones de la Edad de Piedra y las modernas en el este de Asia". Febrero de 2017.
^ Keyser-Tracqui, C; Crubézy, E; Ludes, B (agosto de 2003). "Análisis de ADN nuclear y mitocondrial de una necrópolis de 2000 años de antigüedad en el valle de Egyin Gol de Mongolia". Am. J. Hum. Genet . 73 (2): 247–60. doi :10.1086/377005. PMC 1180365. PMID 12858290 .[ enlace muerto ]
^ abc ИЗУЧЕНИЕ ГЕНЕТИЧЕСКОЙ СТРУКТУРЫ НАРОДОВ ЗАПАДНОГО КАВКАЗА ПО НЫМ О ПОЛИМОРФИЗМЕ Y-ХРОМОСОМЫ, МИТОХОНДРИАЛЬНОЙ ДНК И ALU-ИНСЕРЦИЙ
^ abc Nasidze Iván; Ling, EYS; Quinque, D.; Dupanloup, I.; Cordaux, R.; Rychkov, S.; Naumova, O.; Zhukova, O.; Sarraf-Zadegan, N.; Naderi, GA; Asgary, S.; Sardas, S.; Farhud, DD; Sarkisian, T.; Asadov, C.; Kerimov, A.; Stoneking, M. (2004). "Variación del ADN mitocondrial y del cromosoma Y en el Cáucaso" (PDF) . Anales de genética humana . 68 (parte 3): 205–221. doi :10.1046/j.1529-8817.2004.00092.x. PMID 15180701. S2CID 27204150. Archivado desde el original (PDF) el 18 de enero de 2017. Consultado el 9 de octubre de 2016 .
^ Yunusbayev 2011
^ Haber, Marc; Platt, Daniel E.; Bonab, Maziar Ashrafian; Youhanna, Sonia C.; Soria-Hernanz, David F.; Martínez-Cruz, Begoña; Douaihy, Bouchra; Ghassibe-Sabbagh, Michella; Rafatpanah, Hoshang (2012). "Los grupos étnicos de Afganistán comparten una herencia cromosómica Y estructurada por acontecimientos históricos". MÁS UNO . 7 (3): e34288. Código Bib : 2012PLoSO...734288H. doi : 10.1371/journal.pone.0034288 . ISSN 1932-6203. PMC 3314501 . PMID 22470552.
^ Haber, M; Platt, DE; Ashrafian Bonab, M; Youhanna, SC; Soria-Hernanz, DF; et al. (2012). "Los grupos étnicos de Afganistán comparten una herencia cromosómica Y estructurada por eventos históricos". PLOS ONE . 7 (3): e34288. Bibcode :2012PLoSO...734288H. doi : 10.1371/journal.pone.0034288 . PMC 3314501 . PMID 22470552.
^ abcd Hindu Kush afgano: donde convergen los flujos genéticos del subcontinente euroasiático
^ abcde Introducción de los perfiles de ADN mitocondrial y cromosoma Y argelinos en el panorama del norte de África
^ "Proyecto ADN armenio". Archivado desde el original el 11 de octubre de 2016. Consultado el 8 de octubre de 2016 .
^ abc "¿De dónde vinieron los hombres europeos?" (PDF) . www.jogg.info . 2008 . Consultado el 7 de junio de 2019 .
^ "Comentarios sobre el artículo de Klyosov de 2014 sobre la genealogía del ADN judío, pág. 1 de 2 - ADN levita". Archivado desde el original el 12 de noviembre de 2016. Consultado el 9 de octubre de 2016 .
^ ab Кутуев Ильдус Альбертович. Estructura genética y filología molecular de la escritura
^ ВКЛАД ОТДЕЛЬНЫХ ПОЛЕССКИХ ПОПУЛЯЦИЙ И ПОПУЛЯЦИИ БЕЛОРУССКИХ ТАТАР В ГЕНОФОНД НАСЕЛЕНИЯ БЕЛАРУСИ [1]
^ abc Pericic M, Lauc LB, Klaric IM, et al. (octubre de 2005). "El análisis filogenético de alta resolución del sudeste de Europa rastrea los principales episodios de flujo genético paterno entre las poblaciones eslavas". Mol. Biol. Evol . 22 (10): 1964–75. doi : 10.1093/molbev/msi185 . PMID 15944443.
^ ab Battaglia, Vincenza; Fornarino, Simona; Al-Zahery, Nadia; Olivieri, Anna; Pala, María; Myres, Natalie M; Rey, Roy J; Rootsi, Siiri; et al. (24 de diciembre de 2008). "Evidencia del cromosoma Y de la difusión cultural de la agricultura en el sureste de Europa" (PDF) . Revista europea de genética humana . 17 (6): 820–30. doi :10.1038/ejhg.2008.249. PMC 2947100 . PMID 19107149.
^ Karachanak, Sena; Fornarino, Simona; Grugni, Viola; Semino, Ornella; Toncheva, Draga; Galabov, Ángel; Atanasov, Boris (2009). "Haplogrupos del cromosoma Y en búlgaros". Cuentas rendus de l'Académie bulgare des Sciences . 62 (3): 393–400. ISSN 1310-1331. INIST 21359873.
^ Karachanak S, Grugni V, Fornarino S, Nesheva D, Al-Zahery N, Battaglia V, Carossa V, Yordanov Y, Torroni A, Galabov AS, Toncheva D, Semino O (2013). Pereira LM (ed.). "Diversidad del cromosoma Y en los búlgaros modernos: nuevas pistas sobre su ascendencia". MÁS UNO . 8 (3): e56779. Código Bib : 2013PLoSO...856779K. doi : 10.1371/journal.pone.0056779 . PMC 3590186 . PMID 23483890.
^ "Tabla_Y_Búlgara.xlsx".
^ D. Primorac; et al. (2022). "Herencia genética croata: una historia actualizada del cromosoma Y". Revista Médica Croata . 63 (3): 273–286. doi : 10.3325/cmj.2022.63.273 . PMC 9284021 . PMID 35722696.
^ abcdefghijklm Estructura geográfica del paisaje genético del cromosoma Y del Levante: un contraste entre la costa y el interior
^ Variación del cromosoma Y en la República Checa
^ Altena, E., Smeding, R., van der Gaag, KJ et al. El paisaje cromosómico Y holandés. Eur J Hum Genet 28, 287–299 (2020). https://doi.org/10.1038/s41431-019-0496-0
^ ZONEN VAN ADAM EN NEDERLAND Genetische genealogie: een zoektocht in ons DNA-archief
^ El-Sibai, Mirvat; Platt, Daniel E.; Haber, Marc; Xue, Yalí; Youhanna, Sonia C.; Wells, R. Spencer; Isabel, Hassan; Sanyoura, mayo F.; Harmanani, Haidar (2009). "Estructura geográfica del paisaje genético del cromosoma Y del Levante: un contraste entre la costa y el interior". Anales de genética humana . 73 (6): 568–581. doi :10.1111/j.1469-1809.2009.00538.x. ISSN 1469-1809. PMC 3312577 . PMID 19686289.
^ "Foto" (PNG) . s-media-cache-ak0.pinimg.com . Consultado el 7 de junio de 2019 .
^ Aleros migratorios hacia la región del Báltico
^ "Frecuencias del haplogrupo Y en la población flamenca".
^ Lappalainen, Tuuli; Koivumäki, Satu; Salmela, Elina; Huoponen, Kirsi; Sistonen, Pertti; Savontaus, Marja-Liisa; Lahermo, Päivi (19 de julio de 2006). "Diferencias regionales entre los finlandeses: una perspectiva del cromosoma Y". Gen. 376 (2): 207–215. doi :10.1016/j.gene.2006.03.004. PMID 16644145.
^ Ramos-Luis, E.; Blanco-Verea, A.; Brión, M.; Van Huffel, V.; Carracedo, A.; Sánchez-Diz, P. (diciembre de 2009). "Filogeografía de los linajes masculinos franceses". Forensic Science International: Serie de suplementos genéticos . 2 (1): 439–441. doi : 10.1016/j.fsigss.2009.09.026 . S2CID 85134429.
^ Kayser, Manfred; Lao, Oscar; Anslinger, K; Augustin, Christa; Bargel, G.; Edelmann, J; Elias, Sahar; Heinrich, M; Henke, Jürgen (2005). "La diferenciación genética significativa entre Polonia y Alemania sigue las fronteras políticas actuales, como lo revela el análisis del cromosoma Y". Genética humana . 117 (5): 428–443. doi :10.1007/s00439-005-1333-9. ISSN 0340-6717. PMID 15959808. S2CID 11066186.
^ "Copia archivada" (PDF) . Archivado desde el original (PDF) el 20 de enero de 2017. Consultado el 8 de octubre de 2016 .{{cite web}}: CS1 maint: copia archivada como título ( enlace )(se requiere suscripción)
^ Sanchez, JJ; Børsting, C.; Hernandez, A.; Mengel-Jørgensen, J.; Morling, N. (abril de 2004). "Haplogrupos de SNP del cromosoma Y en daneses, groenlandeses y somalíes". International Congress Series . 1261 : 347–349. doi :10.1016/S0531-5131(03)01635-2.
^ Presencia de tres linajes paternos diferentes entre los indios del norte: un estudio de 560 cromosomas Y (2009)
^ ab Influencias de la historia, la geografía y la religión en la estructura genética: los maronitas en el Líbano
^ Tambets, K; Rootsi, S; Kivisild, T; et al. (abril de 2004). "Las raíces occidentales y orientales de los saami: la historia de los "valores atípicos" genéticos contada por el ADN mitocondrial y los cromosomas Y". Am. J. Hum. Genet . 74 (4): 661–82. doi :10.1086/383203. PMC 1181943 . PMID 15024688.
^ Censo de cromosomas AY de las Islas Británicas
^ Los marcadores uniparentales en Italia revelan una estructura genética sesgada por sexo y diferentes estratos históricos
^ Los patrones clinales de la diversidad cromosómica Y humana en Italia continental y Grecia están dominados por los efectos de deriva y fundador
^ Huellas de acontecimientos históricos olvidados en comunidades de montaña del centro de Italia: una mirada genética
^ Los marcadores uniparentales de la población italiana contemporánea revelan detalles sobre su herencia prerromana
^ La estructura genética de poblaciones aisladas contemporáneas del Tirol del Sur se revela mediante el análisis de los polimorfismos del cromosoma Y, ADNmt y Alu
^ Las migraciones diferenciales griegas y del norte de África a Sicilia están respaldadas por evidencia genética del cromosoma Y
^ Aislamientos en un corredor de migraciones: un análisis de alta resolución de la variación del cromosoma Y en Jordania
^ Вестник Московского университета. Серия XXIII АНТРОПОЛОГИЯ № 1/2014: 96–101СВЯЗЬ ИЗМЕНЧИВОСТИ Y ХРОМОСОМЫ И РОДОВОЙС ТРУКТУРЫ: ГЕНОФОНД СТЕПНОЙ АРИСТОКРАТИИИ ДУХОВЕНСТВА CAZAHOV
^ ab Cromosomas Y en iraníes y tayikos
^ ab Variación del ADNmt y del cromosoma Y en los grupos kurdos
^ El origen dual de los hablantes de tati de Daguestán tal como está escrito en la genealogía de las variantes uniparentales
^ Pliss et al. Linajes del cromosoma Y de los letones en el contexto de la variación genética de la región del Báltico oriental
^ La diversidad del cromosoma Y en el Líbano está estructurada por acontecimientos históricos recientes
^ Linajes paternos en Libia inferidos a partir de haplogrupos del cromosoma Y
^ Evidencia del cromosoma Y de una contribución griega limitada a la población pastún de Pakistán (2006)
^ Historia microfilogeográfica y demográfica de los linajes masculinos portugueses
^ Martínez-Cruz B, Ioana M, Calafell F, Arauna LR, Sanz P, Ionescu R, Boengiu S, Kalaydjieva L, Pamjav H, Makukh H, Plantinga T, van der Meer JW, Comas D, Netea MG (2012). Kivisild T (ed.). "Análisis del cromosoma Y en individuos que llevan el nombre Basarab de la primera dinastía de reyes valacos". MÁS UNO . 7 (7): e41803. Código Bib : 2012PLoSO...741803M. doi : 10.1371/journal.pone.0041803 . PMC 3404992 . PMID 22848614.
^ Diversidad del cromosoma Y de Arabia Saudita y su relación con regiones cercanas.
^ Zgonjanin D, Alghafri R, Antov M, Stojiljković G, Petković S, Vuković R, Drašković D (noviembre de 2017). "Caracterización genética de 27 loci Y-STR con el kit Yfiler ® Plus en la población de Serbia". Internacional de Ciencias Forenses. Genética . 31 : e48-e49. doi :10.1016/j.fsigen.2017.07.013. PMID 28789900.
^ Mihajlovic, Milica; Tanasic, Vanja; Markovic, Milica Keckarevic; Kecmanovic, Miljana; Keckarevic, Dusan (1 de noviembre de 2022). "Distribución de los haplogrupos del cromosoma Y en grupos de población serbios originarios de partes histórica y geográficamente significativas de la península de los Balcanes". Forensic Science International: Genetics . 61 : 102767. doi :10.1016/j.fsigen.2022.102767. ISSN 1872-4973. PMID 36037736. S2CID 251658864.
^ La estructura genética de la población eslovaca revelada por los polimorfismos del cromosoma Y
^ ANALIZA Y-DNK HAPLOTIPOV ESLOVENCEV
^ Adams et al. El legado genético de la diversidad religiosa y la intolerancia: linajes paternos de cristianos, judíos y musulmanes en la península Ibérica
^ Variación del cromosoma Y entre los sudaneses: flujo genético restringido, concordancia con el idioma, la geografía y la historia
^ Karlsson, Andreas O; Wallerstrom, Thomas; Gotherstrom, Anders; Holmlund, Gunilla (2006). "Diversidad del cromosoma Y en Suecia: una perspectiva a largo plazo". Revista Europea de Genética Humana . 14 (8): 963–970. doi : 10.1038/sj.ejhg.5201651 . PMID 16724001.
^ Rootsi S, Magri C, Kivisild T, et al. (julio de 2004). "La filogeografía del haplogrupo I-M170 del cromosoma Y revela dominios distintos del flujo genético prehistórico en Europa". Am. J. Hum. Genet . 75 (1): 128–37. doi :10.1086/422196. PMC 1181996. PMID 15162323 .
^ "Base de datos de haplogrupos suecos (Stats haplopie)". Archivado desde el original el 10 de octubre de 2016. Consultado el 8 de octubre de 2016 .
^ Las distribuciones del cromosoma Y entre las poblaciones del noroeste de China identifican una contribución significativa de los pastores de Asia central y una influencia menor de los euroasiáticos occidentales (2010)
^ Bosch, E.; Calafell, F.; Gonzalez-Neira, A.; Flaiz, C; Mateu, E; Scheil, HG; Huckenbeck, W; Efremovska, L; et al. (2006). "Los linajes paternos y maternos en los Balcanes muestran un paisaje homogéneo a pesar de las barreras lingüísticas, excepto en el caso de los aislados Aromuns". Anales de Genética Humana . 70 (Pt 4): 459–87. doi :10.1111/j.1469-1809.2005.00251.x. PMID 16759179. S2CID 23156886.
^ Lappalainen, T.; Laitinen, V.; Salmela, E.; Andersen, P.; Huoponen, K.; Savontaus, M.-L.; Lahermo, P. (mayo de 2008). "Olas migratorias a la región del Mar Báltico". Ana. Tararear. Genet . 72 (Parte 3): 337–48. doi : 10.1111/j.1469-1809.2007.00429.x . PMID 18294359. S2CID 32079904.
^ ISOGG 2011
^ Cinnioğlu, Cengiz; King, Roy; Kivisild, Toomas; Kalfoğlu, Ersi; Atasoy, Sevil; Cavalleri, Gianpiero L.; Lillie, Anita S.; Roseman, Charles C.; Lin, Alice A. (enero de 2004). "Excavando los estratos de haplotipos del cromosoma Y en Anatolia". Genética humana . 114 (2): 127–148. doi :10.1007/s00439-003-1031-4. ISSN 0340-6717. PMID 14586639. S2CID 10763736.
^ "ISOGG 2017 Haplogrupo I del ADN-Y". isogg.org .
^ ab Caciagli, Laura; Bulayeva, Kazima; Bulayev, Oleg; Bertoncini, Stefania; Taglioli, Luca; Pagani, Luca; Paoli, Giorgio; Tofanelli, Sergio (2009). "El papel clave de la herencia patrilineal en la conformación de la variación genética de los montañeses de Daguestán". Revista de genética humana . 54 (12): 689–694. doi : 10.1038/jhg.2009.94 . ISSN 1434-5161. PMID 19911015.
^ Rootsi, Siiri; Magri, Chiara; Kivisild, Toomas; Benuzzi, Giorgia; Ayuda, Hela; Bermisheva, Marina; Kutuev, Ildus; Barać, Lovorka; Pericic, Marijana (2004). "La filogeografía del haplogrupo I del cromosoma Y revela dominios distintos del flujo de genes prehistóricos en Europa". Revista Estadounidense de Genética Humana . 75 (1): 128-137. doi :10.1086/422196. ISSN 0002-9297. PMC 1181996 . PMID 15162323.
^ Cristofaro J, et al. (octubre de 2013). "Hindu Kush afgano: donde convergen los flujos genéticos del subcontinente euroasiático". PLOS ONE . 8 (10): e76748. Bibcode :2013PLoSO...876748D. doi : 10.1371/journal.pone.0076748 . PMC 3799995 . PMID 24204668.
^ Allentoft, Morten E.; Sikora, Martín; Sjögren, Karl-Göran; Rasmussen, Simón; Rasmussen, Morten; Stenderup, Jesper; Damgaard, Peter B.; Schroeder, Hannes; Ahlström, Torbjörn; Vinner, Lasse; Malaspinas, Anna-Sapfo (junio de 2015). "Genómica de poblaciones de Eurasia de la Edad del Bronce". Naturaleza . 522 (7555): 167–172. Código Bib :2015Natur.522..167A. doi : 10.1038/naturaleza14507. ISSN 1476-4687. PMID 26062507. S2CID 4399103.
^ Malmström, Helena; Gilbert, M. Thomas P.; Thomas, Mark G.; Brandström, Mikael; Storå, Jan; Molnar, Petra; Andersen, Pernille K.; Bendixen, Christian; Holmlund, Gunilla; Götherström, Anders; Willerslev, Eske (3 de noviembre de 2009). "El ADN antiguo revela una falta de continuidad entre los cazadores-recolectores neolíticos y los escandinavos contemporáneos". Current Biology . 19 (20): 1758–1762. doi : 10.1016/j.cub.2009.09.017 . ISSN 0960-9822. PMID 19781941.
^ Karlsson, Andreas O.; Wallerström, Thomas; Götherström, Anders; Holmlund, Gunilla (agosto de 2006). "Diversidad del cromosoma Y en Suecia: una perspectiva a largo plazo". Revista Europea de Genética Humana . 14 (8): 963–970. doi : 10.1038/sj.ejhg.5201651 . ISSN 1476-5438. PMID 16724001.
^ abc Rootsi; et al. "La filogeografía del haplogrupo I del cromosoma Y revela dominios distintos del flujo genético prehistórico en Europa, figura 1" (PDF) . Archivado desde el original (PDF) el 3 de marzo de 2007.
^ Marjanovic D, Fornarino S, Montagna S, et al. (noviembre de 2005). "El poblamiento de la Bosnia-Herzegovina moderna: haplogrupos del cromosoma Y en los tres grupos étnicos principales". Ann. Hum. Genet . 69 (parte 6): 757–63. doi :10.1111/j.1529-8817.2005.00190.x. PMID 16266413. S2CID 36632274.[ enlace muerto ]
^ Om Utevska (2017). Genofond українців за різними системами генетичних маркерів: походження і місце на європейському генетичному просторі [ El acervo genético de los ucranianos revelado por diferentes sistemas de marcadores genéticos: el origen y la presencia en Europa ] (PhD) (en ucraniano). Centro Nacional de Investigación de Medicina Radiológica de la Academia Nacional de Ciencias de Ucrania . págs. 219–226, 302.
^ McEvoy y Bradley, Brian P y Daniel G (2010). Celtic from the West Capítulo 5: Irish Genetics and Celts . Oxbow Books, Oxford, Reino Unido. pág. 117. ISBN978-1-84217-410-4.
^ Haber, Marc; Platt, Daniel E.; Ashrafian Bonab, Maziar; Youhanna, Sonia C.; Soria-Hernanz, David F.; Martínez-Cruz, Begoña; Douaihy, Bouchra; Ghassibe-Sabbagh, Michella; Rafatpanah, Hoshang; Ghanbari, Mohsen; Ballena, Juan; Balanovsky, Oleg; Wells, R. Spencer; Comas, David; Tyler-Smith, Chris; Zalloua, Pierre A. (2012). "Los grupos étnicos de Afganistán comparten una herencia cromosómica Y estructurada por acontecimientos históricos". MÁS UNO . 7 (3): e34288. Código Bib : 2012PLoSO...734288H. doi : 10.1371/journal.pone.0034288 . PMC 3314501 . Número de modelo: PMID22470552.
^ Grasgruber, Pavel; Popović, Stevo; Bokuvka, Dominik; Davidović, Ivan; Hřebíčková, Sylva; Ingrová, Pavlína; Potpara, Predrag; Precio, Stipan; Stračárová, Nikola (2017). "Las montañas de los gigantes: un estudio antropométrico de jóvenes varones en Bosnia y Herzegovina". Ciencia abierta de la Royal Society . 4 (4): 161054. Código bibliográfico : 2017RSOS....461054G. doi :10.1098/rsos.161054. PMC 5414258 . PMID 28484621.
^ Grasgruber, P.; Cacek, J.; Kalina, T.; Sebera, M. (1 de diciembre de 2014). "El papel de la nutrición y la genética como determinantes clave de la tendencia positiva de la altura". Economía y biología humana . 15 : 81–100. doi : 10.1016/j.ehb.2014.07.002 . ISSN 1570-677X. PMID 25190282.
Enlaces externos
Wikimedia Commons alberga una categoría multimedia sobre Haplogrupo I del ADN-Y .
Árbol filogenético y mapas de distribución
Haplogrupo I-M170 de ADN-Y y sus subclados de la Sociedad Internacional de Genealogía Genética (ISOGG) de 2013
Filogeografía del haplogrupo I del cromosoma Y
Distribuciones de frecuencias del haplogrupo I del ADN-Y y sus subclados (con videotutorial)
Frecuencia y varianza de I1b (ahora considerada I2a2-M26)
Mapa de 'I1a' (ahora considerado I-M253)
Mapa de 'I1b' Archivado el 1 de mayo de 2015 en Wayback Machine (ahora considerado I2a-P37.2)
Mapa de 'I1c' Archivado el 16 de agosto de 2008 en Wayback Machine (ahora considerado I2b-M223)
Se le cambió el nombre a Árbol del Haplogrupo I (K. Nordtvedt 2011).
Proyectos
Proyecto I en FTDNA
Proyecto I1 en FTDNA
Proyecto I2* en FTDNA
Proyecto I2a en FTDNA
Proyecto I2b en FTDNA
Proyecto I2b2 L38+ en FTDNA
El Proyecto Genealógico Escandinavo de ADN-Y en FTDNA
El Proyecto Genealógico de Finlandia en FTDNA
Otro
Estudio del haplogrupo I del cromosoma Y y de los haplotipos modales
El Consorcio del Cromosoma Y (YCC)
Ejemplos de haplotipos del grupo I1* "y"
Página del haplogrupo I del YCC: I1a (ahora considerado I-M253), I1b (ahora considerado I-P37.2) e I1c (ahora considerado I-M223)
Predictor del subclade Haplo-I Archivado el 2 de abril de 2022 en Wayback Machine