R1b tiene dos ramas principales: R1b1-L754 y R1b2-PH155. R1b1-L754 tiene dos subclados principales: R1b1a1b-M269, que predomina en Europa occidental, y R1b1b-V88, que hoy es común en partes de África central. La otra rama, R1b2-PH155, es tan rara y está tan ampliamente dispersa que es difícil sacar conclusiones sobre sus orígenes. Se ha encontrado en Bahréin , India , Nepal , Bután , Ladakh , Tayikistán , Turquía y China occidental .
Según estudios de ADN antiguo, la mayoría de los linajes R1a y R1b se habrían expandido desde la estepa póntica junto con las lenguas indoeuropeas . [2] [3] [4] [5] [6]
Origen y dispersión
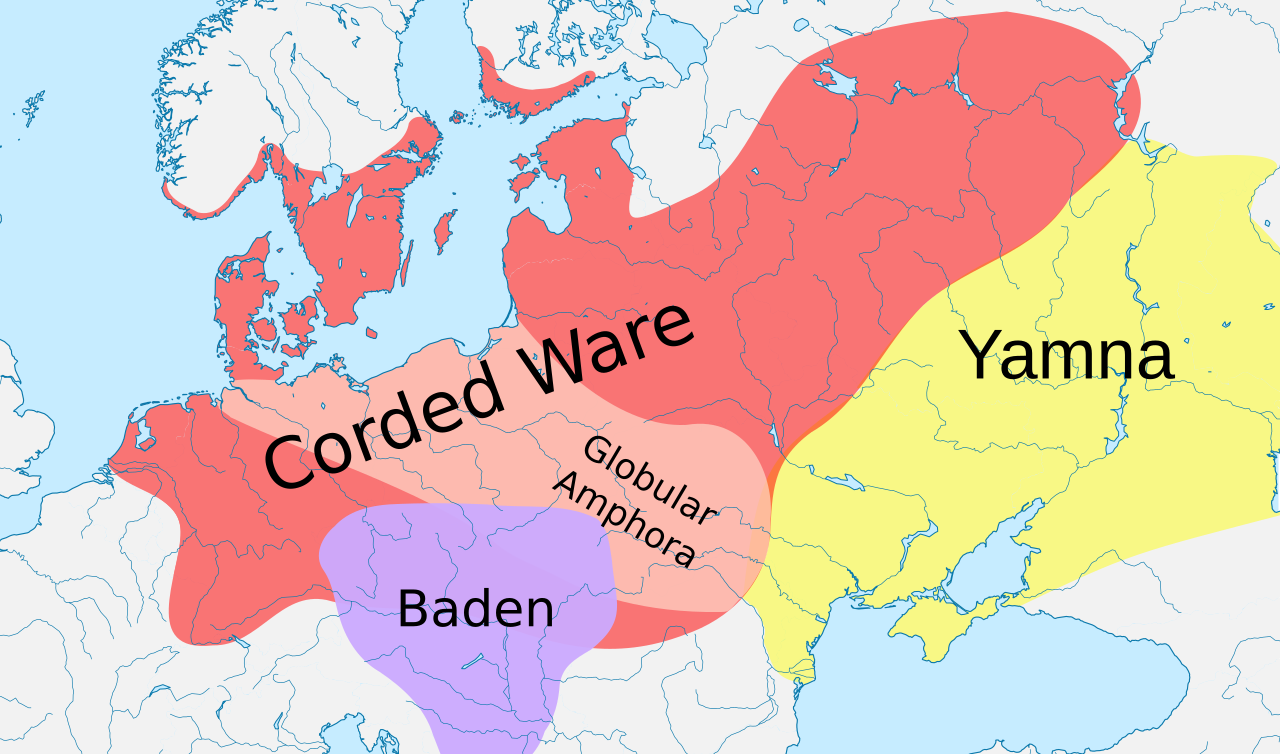
Estudios genéticos realizados desde 2015 han revelado que la cultura Yamna , que se cree que hablaba alguna etapa del protoindoeuropeo , era portadora del R1b-L23.
La edad de R1 fue estimada por Tatiana Karafet et al. (2008) entre 12.500 y 25.700 BP , y lo más probable es que haya ocurrido hace unos 18.500 años. [7] Dado que el ejemplo más antiguo conocido ha sido datado en alrededor de 14.000 BP, y pertenece a R1b1 (R-L754), [1] R1b debe haber surgido relativamente pronto después del surgimiento de R1.
Entre los primeros restos humanos que portaban R1b se encuentran:
Varios varones mesolíticos y neolíticos enterrados en Deriivka y Vasil'evka en la actual Ucrania entre 9500 y 7000 años antes del presente eran portadores de R1b1a. [12] Estos individuos eran en su mayoría de ascendencia EHG, con una importante mezcla de WHG. [13]
Un hombre de WHG enterrado en Ostrovul Corbuli, Rumania , alrededor del año 8700 a. C., portaba R1b1c. [14] [15] [16]
Un hombre enterrado en Lepenski Vir , Serbia, entre 8200 y 7900 años antes del presente, portaba R1b1a. [17]
Un EHG enterrado cerca de Samara , Rusia, hace 7500 años contenía R1b1a1a. [18]
Un hombre del Eneolítico enterrado en Khvalynsk , Rusia, entre 7200 y 6000 años antes del presente, portaba R1b1a. [19]
Se descubrió que un hombre neolítico enterrado en Els Trocs , España, entre 7178 y 7066 a. C., que podría haber pertenecido a la cultura Epicárdica , [20] era portador de R1b1. [21] [22] [23]
Un hombre del Calcolítico Tardío enterrado en Smyadovo , Bulgaria, alrededor del año 6500 a. C., portaba R1b1a. [24]
Un hombre de la Edad del Cobre Temprano enterrado en Cannas di Sotto, Carbonia, Cerdeña alrededor del año 6450 a. C., portaba R1b1b2. [25]
Un hombre del grupo Baalberge en Europa Central enterrado alrededor del año 5600 a. C. portaba R1b1a. [26]
Un hombre de la cultura Botai en Asia Central enterrado alrededor del año 5500 a. C. portaba R1b1a1 (R1b-M478). [27]
Se encontró que 7 machos de la cultura Yamnaya que fueron analizados pertenecían al subclado M269 del haplogrupo R1b. [28]
R1b es un subclado dentro del "macro- haplogrupo " K (M9), el grupo más común de líneas masculinas humanas fuera de África. Se cree que K se originó en Asia (como es el caso de un haplogrupo ancestral aún más temprano, F (F-M89). Karafet T. et al. (2014) sugirieron que un "rápido proceso de diversificación de K-M526 probablemente ocurrió en el sudeste asiático , con posteriores expansiones hacia el oeste de los ancestros de los haplogrupos R y Q ". [29] Sin embargo, el ejemplo más antiguo de R* se ha encontrado en una muestra de Eurasia del Norte Antiguo de Siberia ( niño de Mal'ta , hace 24.000 años), y su precursor P1 se ha encontrado en otra muestra de Eurasia del Norte Antiguo de Siberia del norte ( Yana RHS ) que data de hace c. 31.600 años. [30]
Tres estudios genéticos realizados en 2015 dieron apoyo a la hipótesis Kurgan de Marija Gimbutas sobre la patria protoindoeuropea . Según esos estudios, los haplogrupos R1b-M269 y R1a, ahora los más comunes en Europa (R1a también es común en el sur de Asia) se habrían expandido desde la estepa euroasiática occidental, junto con las lenguas indoeuropeas ; también detectaron un componente autosómico presente en los europeos modernos que no estaba presente en los europeos neolíticos , que se habría introducido con los linajes paternos R1b y R1a, así como con las lenguas indoeuropeas. [2] [3] [4]
El análisis del ADN-Y antiguo de los restos de los asentamientos de la cultura de cerámica lineal del Neolítico temprano en Europa central y del norte aún no ha encontrado varones pertenecientes al haplogrupo R1b-M269. [31] [32] Olalde et al. (2017) rastrean la propagación del haplogrupo R1b-M269 en Europa occidental, particularmente en Gran Bretaña, hasta la propagación de la cultura Beaker , con una aparición repentina de muchos haplogrupos R1b-M269 en Europa occidental ca. 5000-4500 años AP durante la Edad del Bronce temprana. [33]
Estructura
Filogenia externa de R1b
El haplogrupo más amplio R (M207) es un subclado primario del haplogrupo P1 (M45), que a su vez es una rama primaria de P (P295), que también se conoce como haplogrupo K2b2. Por lo tanto, R-M207 es una rama secundaria de K2b (P331) y un descendiente directo de K2 (M526).
No se han notificado casos confirmados de R1b* (R-M343*), es decir, R1b (xR1b1, R1b2), también conocido como R-M343 (xL754, PH155), en la literatura revisada por pares .
R-M343 (M73, M269, V88)
En las primeras investigaciones, debido a que R-M269, R-M73 y R-V88 son por lejos las formas más comunes de R1b, a veces se asumió que los ejemplos de R1b (xM73, xM269) significaban ejemplos basales de "R1b*". [35] Sin embargo, si bien el paragrupo R-M343 (xM73, M269, V88) es raro, no excluye la pertenencia a subclados relativamente basales, raros y/o descubiertos posteriormente, de R1b, como R-L278* (R1b*), R-L389* (R1b1a*), R-P297* (R1b1a1*), R-V1636 (R1b1a2) o R-PH155 (R1b2).
Se cree que la población que tiene la mayor proporción de R-M343 (xM73, M269, V88) son los kurdos del sureste de Kazajstán con un 13%. [41] Sin embargo, más recientemente, un estudio a gran escala sobre la variación del cromosoma Y en Irán reveló que la presencia de R-M343 (xV88, M73, M269) llega a alcanzar un 4,3% entre las subpoblaciones iraníes. [42]
Sigue existiendo la posibilidad de que algunos, o incluso la mayoría de estos casos, puedan ser R-L278* (R1b*), R-L389* (R1b1a*), R-P297* (R1b1a1*), R-V1636 (R1b1a2), R-PH155 (R1b2), R1b* (R-M343*), R1a* (R-M420*), una rama no documentada de R1 (R-M173), y/o retromutaciones de un marcador, de un estado ancestral positivo a uno negativo, [43] y, por lo tanto, constituir subclados no documentados de R1b.
En Cruciani et al. (2010) [44] se puede encontrar una recopilación de estudios previos sobre la distribución de R1b. Se resume en la siguiente tabla. (Cruciani no incluyó algunos estudios que sugerían frecuencias incluso más altas de R1b1a1b [R-M269] en algunas partes de Europa occidental).
R1b (R-L278)
El gen R-L278 entre los hombres modernos pertenece a los subclados R-L754 y R-PH155, aunque es posible que exista algún gen R-L278* muy raro, ya que no se han analizado todos los ejemplos para ambas ramas. También pueden existir ejemplos en el ADN antiguo, aunque debido a la mala calidad, a menudo es imposible determinar si los antiguos portaban o no las mutaciones que definen los subclados.
Algunos ejemplos descritos en artículos más antiguos, por ejemplo dos encontrados en Turquía, [34] ahora se cree que están principalmente en el subclado más recientemente descubierto R1b1b (R-V88). La mayoría de los ejemplos de R1b, por lo tanto, caen en los subclados R1b1b (R-V88) o R1b1a (R-P297). Cruciani et al. en el gran estudio de 2010 encontraron 3 casos entre 1173 italianos, 1 de 328 asiáticos occidentales y 1 de 156 asiáticos orientales. [44] Varzari encontró 3 casos en Ucrania , en un estudio de 322 personas de la región de los Montes Dniéster - Cárpatos , que fueron P25 positivos, pero M269 negativos. [45] Los casos de estudios más antiguos son principalmente de África, Medio Oriente o el Mediterráneo, y se analizan a continuación como casos probables de R1b1b (R-V88).
R1b1 (R-L754)
R-L754 contiene la gran mayoría de R1b. El único ejemplo conocido de R-L754* (xL389, V88) es también el individuo más antiguo conocido que portaba R1b: " Villabruna 1 ", que vivió hace unos 14.000 años (al noreste de Italia). Villabruna 1 pertenecía a la cultura del Epigravetiense .
R1b1a (R-L389)
R-L389, también conocido como R1b1a (L388/PF6468, L389/PF6531), contiene el subclado muy común R-P297 y el subclado poco común R-V1636. Se desconoce si todos los R-L389* (xP297) informados anteriormente pertenecen o no a R-V1636.
R1b1a1 (R-P297)
El marcador SNP P297 fue reconocido en 2008 como ancestral de los subclados significativos M73 y M269, combinándolos en un solo grupo. [7] Se le había dado el nombre filogenético R1b1a1a (y, anteriormente, R1b1a).
La mayoría de los R1b euroasiáticos pertenecen a este subclado, lo que representa una población moderna muy grande. Aunque el P297 en sí no ha sido muy estudiado todavía, la misma población ha sido relativamente bien estudiada en términos de otros marcadores. Por lo tanto, la ramificación dentro de este clado se puede explicar con bastante detalle a continuación.
R1b1a1a (R-M73)
Malyarchuk et al. (2011) encontraron R-M73 en el 13,2% (5/38) de los shors, el 11,4% (5/44) de los teleuts, el 3,3% (2/60) de los kalmyks, el 3,1% (2/64) de los khakassianos, el 1,9% (2/108) de los tuvinianos y el 1,1% (1/89) de los altaianos. [46] Los kalmyks, tuvinianos y altaianos pertenecen a un grupo Y-STR marcado por DYS390=19, DYS389=14-16 (o 14-15 en el caso del individuo altaiano) y DYS385=13-13.
Dulik et al. (2012) encontraron R-M73 en el 35,3% (6/17) de una muestra de los Kumandin de la República de Altai en Rusia. [47] Tres de estos seis Kumandin comparten un haplotipo Y-STR idéntico de 15 loci, y otros dos difieren solo en el locus DYS458, teniendo DYS458=18 en lugar de DYS458=17. Este par de haplotipos R-M73 de Kumandin se asemeja a los haplotipos de dos Kalmyks, dos Tuvinianos y un Altaiano cuyo Y-ADN ha sido analizado por Malyarchuk et al. (2011). El Kumandin R-M73 restante tiene un haplotipo Y-STR que es marcadamente diferente de los haplotipos de los otros Kumandin R-M73, asemejándose en cambio a los haplotipos de cinco Shors, cinco Teleuts y dos Khakassianos. [46]
Aunque las primeras investigaciones sobre R-M73 afirmaban que estaba significativamente representado entre los hazara de Afganistán y los bashkires de los montes Urales, esto aparentemente ha sido revocado. Por ejemplo, el material de apoyo de un estudio de 2010 de Behar et al. sugirió que Sengupta et al. (2006) podrían haber identificado erróneamente a los individuos hazara, que en cambio pertenecían a "PQR2" en lugar de "R(xR1a)". [48] [41] [49] Sin embargo, la asignación del ADN-Y de estos hazaras a la categoría "PQR2" por Behar et al. (2010) probablemente se pueda atribuir al hábito que fue popular durante un tiempo de etiquetar a R-M269 como "R1b" o "R(xR1a)", y colocar a cualquier miembro de R-M343 (xM269) en una categoría polifilética, general, "R*" o "P". Myres et al. (2011), Di Cristofaro et al. (2013), y Lippold et al. (2014) coinciden en que el ADN-Y del 32% (8/25) de la muestra HGDP de hazara paquistaníes debería pertenecer al haplogrupo R-M478/M73. [41] [50] [51] Del mismo modo, se ha descubierto que la mayoría de los varones baskires pertenecen a U-152 (R1b1a1a2a1a2b) y algunos, principalmente del sureste de Bashkortostán, pertenecían al haplogrupo Q-M25 (Q1a1b) en lugar de R1b; en contra de esto, Myres et al. (2011) encontraron una alta frecuencia de R-M73 entre su muestra de baskires del sureste de Bashkortostán (77/329 = 23,4% R1b-M73), de acuerdo con el estudio anterior de baskires. [41] Además de la alta frecuencia de R-M73 en los bashkires del sureste, Myres et al. También se informó del hallazgo de R-M73 en las siguientes muestras: 10,3% (14/136) de los balkarios del noroeste del Cáucaso, 9,4% (8/85) de las muestras de HGDP del norte de Pakistán (estos son los hazaras paquistaníes antes mencionados), 5,8% (4/69) de los karachái del noroeste del Cáucaso, 2,6% (1/39) de los tártaros de Baskortostán, 1,9% (1/54) de los baskires del suroeste de Baskortostán, 1,5% (1/67) de los megrelos del sur del Cáucaso, 1,4% (1/70) de los baskires del norte de Baskortostán, 1,3% (1/80) de los tártaros de Kazán, 1,1% (1/89) de una muestra de Capadocia, Turquía, 0,7% (1/141) de los kabardianos del noroeste del Cáucaso, el 0,6% (3/522) de un grupo de muestras de Turquía y el 0,38% (1/263) de rusos de Rusia Central. [41]
Además de los hazaras paquistaníes antes mencionados, Di Cristofaro et al. (2013) encontraron R-M478/M73 en el 11,1% (2/18) de los mongoles de Mongolia central, el 5,0% (1/20) de los kirguisos del suroeste de Kirguistán, el 4,3% (1/23) de los mongoles del sureste de Mongolia, el 4,3% (4/94) de los uzbekos de Jawzjan, Afganistán, el 3,7% (1/27) de los iraníes de Gilan , el 2,5% (1/40) de los kirguisos del centro de Kirguistán, el 2,1% (2/97) de los mongoles del noroeste de Mongolia y el 1,4% (1/74) de los turcomanos de Jawzjan, Afganistán. [50] Los mongoles, así como el individuo del suroeste de Kirguistán, el individuo de Gilan y uno de los uzbekos de Jawzjan pertenecen al mismo grupo de haplotipos Y-STR que cinco de los seis miembros kumandin de R-M73 estudiados por Dulik et al. (2012). El valor Y-STR más distintivo de este grupo es DYS390=19. [41]
Karafet et al. (2018) encontraron R-M73 en el 37,5% (15/40) de una muestra de teleuts de Bekovo, óblast de Kemerovo, 4,5% (3/66) de una muestra de uigures de la Región Autónoma Uigur de Xinjiang, 3,4% (1/29) de una muestra de kazajos de Kazajstán, 2,3% (3/129) de una muestra de selkups, 2,3% (1/44) de una muestra de turcomanos de Turkmenistán y 0,7% (1/136) de una muestra de iraníes de Irán. [52] Cuatro de estos individuos (uno de los teleuts, uno de los uigures, el kazajo y el iraní) parecen pertenecer al grupo mencionado anteriormente marcado por DYS390=19 (el grupo R-M73 kumandin-mongol); Los teleut y los uigures también comparten los valores modales en los loci DYS385 y DYS389. El iraní difiere del modal para este grupo al tener 13-16 (o 13-29) en DYS389 en lugar de 14-16 (o 14-30). El kazajo difiere del modal al tener 13-14 en DYS385 en lugar de 13-13. Los otros catorce teleuts y los tres selkups parecen pertenecer al grupo teleut-shor-khakassian R-M73 del conjunto de datos de Malyarchuk et al. (2011); Este grupo tiene los valores modales de DYS390=22 (pero 21 en el caso de dos teleuts y un khakassiano), DYS385=13-16 y DYS389=13-17 (o 13-30, pero 14-31 en el caso de un selkup).
Un artículo kazajo publicado en 2017 encontró el haplogrupo R1b-M478 Y-DNA en el 3,17% (41/1294) de una muestra de kazajos de Kazajstán, y este haplogrupo se observó con una frecuencia mayor que la media entre los miembros de las tribus Qypshaq (12/29 = 41,4%), Ysty (6/57 = 10,5%), Qongyrat (8/95 = 8,4%), Oshaqty (2/29 = 6,9%), Kerey (1/28 = 3,6%) y Jetyru (3/86 = 3,5%) . [ 53] Un artículo chino publicado en 2018 encontró el haplogrupo R1b-M478 Y-DNA en el 9,2% (7/76) de una muestra de uigures dolan del municipio de Horiqol, condado de Awat , Xinjiang. [54]
R1b1a1b (R-M269)
El R-M269, o R1b1a1b (a partir de 2018), entre otros nombres, [55] es actualmente el linaje de ADN-Y más común en los varones europeos. Se estima que lo portan unos 110 millones de varones en Europa. [56]
Distribución de frecuencias espaciales proyectadas dentro de Europa del haplogrupo R-M269. [56]
El R-M269 ha recibido un gran interés científico y popular debido a su posible conexión con la expansión indoeuropea en Europa. En concreto, se ha descubierto que el subclado R-Z2103 es frecuente en el ADN antiguo asociado con la cultura Yamna . [2] Se determinó que los siete individuos de un grupo pertenecían al subclado R1b-M269. [2]
Investigaciones más antiguas, publicadas antes de que los investigadores pudieran estudiar el ADN de los restos antiguos, propusieron que R-M269 probablemente se originó en Asia occidental y estuvo presente en Europa en el período Neolítico. [36] [41] [57] [58] Pero los resultados basados en ADN antiguo real notaron que hubo una escasez de R-M269 en Europa antes de la Edad del Bronce, [2] y la distribución de subclados dentro de Europa se debe sustancialmente a las diversas migraciones de la Edad del Bronce y la Edad del Hierro . Asimismo, las muestras más antiguas clasificadas como pertenecientes a R-M269 se han encontrado en Europa del Este y la estepa póntico-caspia, no en Asia occidental. Las poblaciones de Europa occidental se dividen entre los subclados R-P312/S116 y R-U106/S21 de R-M412 (R-L51).
La distribución de R-M269 en Europa aumenta en frecuencia de este a oeste. Alcanza su pico a nivel nacional en Gales a una tasa del 92%, del 82% en Irlanda , del 70% en Escocia , del 68% en España , del 60% en Francia (76% en Normandía ), alrededor del 60% en Portugal , [41] 50% en Alemania , 50% en los Países Bajos , 47% en Italia , [59] 45% en el este de Inglaterra y 42% en Islandia . R-M269 alcanza niveles tan altos como el 95% en partes de Irlanda. También se ha encontrado en frecuencias más bajas en toda Eurasia central , [60] pero con una frecuencia relativamente alta entre los bashkires de la región de Perm (84,0%). [61] Este marcador está presente en China e India en frecuencias de menos del uno por ciento. En el norte de África y las islas adyacentes, mientras que R-V88 (R1b1b) está más fuertemente representado, R-M269 parece haber estado presente desde la antigüedad. R-M269 se ha encontrado, por ejemplo, a una tasa de ~44% entre restos que datan de los siglos XI al XIII en Punta Azul , en las Islas Canarias . Estos restos se han relacionado con los bimbaches (o bimape), un subgrupo de los guanches. [62] En los machos vivos, alcanza su punto máximo en partes del norte de África, especialmente Argelia , a una tasa del 10%. [63] En el África subsahariana, R-M269 parece alcanzar su punto máximo en Namibia , a una tasa del 8% entre los machos herero . [64] En Asia occidental, R-M269 se ha informado en el 40% de los machos armenios y en más del 35% en los machos turcomanos . [65] [66] (La siguiente tabla enumera con más detalle las frecuencias de M269 en regiones de Asia, Europa y África).
Aparte del R-M269* basal que no ha divergido, existen (a fecha de 2017) dos ramas principales del R-M269:
R-L23 (R1b1a1b1; L23/PF6534/S141) y
R-PF7558 (R1b1a1b2; PF7558/PF7562.)
Se ha informado de R-L23 (Z2105/Z2103; también conocido como R1b1a1b1) entre los pueblos de Idel-Ural (por Trofimova et al. 2015): 21 de 58 (36,2 %) de los baskires del distrito de Burzyansky , 11 de 52 (21,2 %) de los udmurtos , 4 de 50 (8 %) de los komi , 4 de 59 (6,8 %) de los mordvinos , 2 de 53 (3,8 %) de los besermios y 1 de 43 (2,3 %) de los chuvasos eran R1b-L23. [67]
Los subclados dentro del paragrupo R-M269(xL23) – es decir, R-M269* y/o R-PF7558 – parecen encontrarse en su frecuencia más alta en los Balcanes centrales , especialmente Kosovo con 7,9%, Macedonia del Norte 5,1% y Serbia 4,4%. [41] A diferencia de la mayoría de las otras áreas con porcentajes significativos de R-L23, Kosovo , Polonia y los bashkires del sureste de Bashkortostán son notables por tener un alto porcentaje de R-L23 (xM412) – en tasas de 11,4% (Kosovo), 2,4% (Polonia) y 2,4% sureste de Bashkortostán. [41] (Esta población bashkir también es notable por su alto nivel de R-M73 (R1b1a1a1), con un 23,4%. [41] ) Cinco individuos de 110 evaluados en el valle de Ararat de Armenia pertenecían a R-M269(xL23) y 36 a R-L23*, y ninguno pertenecía a subclados conocidos de L23. [68]
En 2009, el ADN extraído de los huesos del fémur de 6 esqueletos en un lugar de enterramiento de principios de la Edad Media en Ergolding (Baviera, Alemania) que data de alrededor del año 670 d. C. arrojó los siguientes resultados: se encontró que 4 eran del haplogrupo R1b con las coincidencias más cercanas en las poblaciones modernas de Alemania, Irlanda y los EE. UU., mientras que 2 estaban en el haplogrupo G2a . [69]
A continuación se ofrece un resumen de la mayoría de los estudios que probaron específicamente M269, mostrando su distribución (como porcentaje de la población total) en Europa, el norte de África , Oriente Medio y Asia central hasta China y Nepal .
La filogenia de R-M269 según ISOGG 2017:
R1b1b (R-V88)
R1b1b (PF6279/V88; anteriormente R1b1a2) se define por la presencia del marcador SNP V88, cuyo descubrimiento fue anunciado en 2010 por Cruciani et al. [44] Aparte de los individuos en el sur de Europa y Asia occidental, la mayoría de R-V88 se encontró en el Sahel, especialmente entre poblaciones que hablan lenguas afroasiáticas de la rama chádica .
Basándose en un análisis filogenético detallado, D'Atanasio et al. (2018) propusieron que R1b-V88 se originó en Europa hace unos 12.000 años y cruzó al norte de África hace entre 8000 y 7000 años, durante el período del " Sahara Verde ". R1b-V1589, el subclado principal dentro de R1b-V88, experimentó una mayor expansión hace unos 5500 años, probablemente en la región de la cuenca del lago Chad, desde donde algunas líneas volvieron a cruzar el Sahara hasta el norte de África. [70]
Marcus et al. (2020) aportan pruebas sólidas de este modelo propuesto de movimiento transsahariano de norte a sur: los primeros haplogrupos basales R1b-V88 se encuentran en varios cazadores-recolectores de Europa del Este hace cerca de 11.000 años. Luego, el haplogrupo aparentemente se extendió con la expansión de los agricultores neolíticos , que establecieron la agricultura en el Mediterráneo occidental alrededor de 7500 AP. Los haplogrupos R1b-V88 se identificaron en individuos neolíticos antiguos en Alemania, Italia central, Iberia y, con una frecuencia particularmente alta, en Cerdeña. Una parte de la rama que conduce a los haplogrupos africanos actuales (V2197) ya se había derivado de individuos europeos neolíticos de España y Cerdeña, lo que proporciona más apoyo a un movimiento transsahariano de norte a sur. [71] [72] [73] También se han identificado ascendencia autosómica europea, haplogrupos de ADNmt y alelos de persistencia de la lactasa en poblaciones africanas que portan R1b-V88 con una alta frecuencia, como los fulani y los toubou . [74] [75] [72] [76] La presencia de agricultores neolíticos europeos en África está atestiguada además por muestras de Marruecos que datan de aproximadamente el 5400 a. C. en adelante. [77] [78]
Estudios realizados entre 2005 y 2008 informaron de niveles elevados de "R1b*" en Jordania , Egipto y Sudán . [79] [64] [80] [nota 1] Investigaciones posteriores de Myres et al. (2011) indican que las muestras en cuestión probablemente pertenecen al subclado R-V88. Según Myres et al. (2011), esto puede explicarse por una migración de retorno desde Asia a África por parte de personas portadoras de R1b. [41] [nota 2]
A diferencia de otros estudios, Shriner y Rotimi (2018) asociaron la introducción de R1b en Chad con los movimientos más recientes de los árabes baggaras . [81]
Dos ramas del R-V88, R-M18 y R-V35, se encuentran casi exclusivamente en la isla de Cerdeña .
Como se puede ver en la tabla de datos anterior, el R-V88 se encuentra en el norte de Camerún , en el centro-oeste de África, con una frecuencia muy alta, donde se considera que es causado por un movimiento preislámico de personas procedentes de Eurasia . [64] [82]
R1b1b1 (R-M18)
R1b1b1 es un subclado de R-V88, que se define por la presencia del marcador SNP M18. [7]
Se ha encontrado solo en frecuencias bajas en muestras de Cerdeña [60] [83] y Líbano . [84]
R1b2 (R-PH155)
R1b2 es extremadamente raro y se define por la presencia de PH155. [36] Se han encontrado machos vivos portadores de subclados de R-PH155 en Bahréin , Bután , Ladakh , Tayikistán , Turquía, Xinjiang y Yunnan . ISOGG (2022) cita dos ramas principales: R-M335 (R1b2a) y R-PH200 (R1b2b).
El SNP definitorio de R1b2a, M335, se documentó por primera vez en 2004, cuando se descubrió un ejemplo en Turquía, aunque en ese momento se clasificó como R1b4. [34] Se han informado otros ejemplos de R-M335 en una muestra de Hui de Yunnan , China [85] y en una muestra de personas de Ladakh , India. [86]
En la cultura popular
Bryan Sykes , en su libro de 2006 Blood of the Isles , da a los miembros -y al supuesto patriarca fundador- de R1b el nombre " Oisín ".
Stephen Oppenheimer , en su libro de 2007 Origins of the British , le da al patriarca R1b el nombre vasco "Ruisko" en honor a lo que Oppenheimer creía que era el origen ibérico de R1b.
Un cineasta llamado Artem Lukichev creó (alrededor de 2009) una película animada de 14 minutos basada en una epopeya bashkir de los montes Urales , relacionando la epopeya con el surgimiento y la expansión geográfica de R1a y R1b. [87]
Las pruebas de ADN que ayudaron a la identificación del zar Nicolás II de Rusia encontraron que pertenecía a R1b. [88] Esto puede sugerir que los zares posteriores de la Casa de Romanov , descendientes en la línea masculina de la Casa de Holstein-Gottorp (que se originó en Schleswig-Holstein ) también son miembros de R1b.
Personas notables
Spytihněv I, duque de Bohemia , las pruebas de ADN realizadas a sus restos sugieren que su haplogrupo Y era R1b. [89]
La Casa de Borbón , que ha gobernado como reyes en Francia, España y otros países europeos, tiene el haplogrupo R1b1b. [90]
Las pruebas de ADN realizadas a varias momias de la XVIII dinastía de Egipto revelaron el haplogrupo R1b. La momia de Tutankamón tenía el haplogrupo Y R1b y el haplogrupo K de ADNmt . Heredó este haplogrupo Y de su padre, la momia KV55 que muchos creen que fue Akenatón , y de su abuelo, Amenhotep III , cuya momia fue encontrada sepultada en KV35 con numerosos familiares. [91] [92]
Salud
Los estudios han demostrado que el haplogrupo R1b podría tener un efecto protector sobre el sistema inmunológico. [93] Sin embargo, estudios posteriores han confirmado que el cromosoma Y tiene un efecto muy limitado sobre la enfermedad de la arteria coronaria (EAC), por ejemplo, y que el vínculo previamente supuesto entre los haplogrupos del cromosoma Y y la salud está lejos de establecerse científicamente. [94]
Véase también
Wikimedia Commons alberga una categoría multimedia sobre Haplogrupo R1b del ADN-Y .
^ Flores et al. (2005) encontraron que 20 de los 146 hombres examinados (13,7%) –incluyendo 20 de los 45 hombres examinados de la zona del Mar Muerto de Jordania– dieron positivo para M173 (R1), y negativo para los marcadores R1a SRY10831.2 y M17, así como para P25 (que más tarde se descubrió que era un marcador poco fiable para R1b1). Myres et al. (2011) [41] indica que son en su mayoría R-V88 (más tarde conocido como R1b1b). Wood et al. (2005) también informaron de dos casos egipcios de R1* (R-M173*) que dieron negativo para SRY10831 (R1a1) y el marcador poco fiable R1b1 P25, de una muestra de 1.122 hombres de países africanos, incluidos 92 de Egipto. Hassan et al. (2008) encontraron un número igualmente sorprendente de 14 de 26 (54%) de los fula sudaneses que eran M173+ y P25-.
^ Myres et al. (2011): "La detección de V88 en Irán, Palestina y especialmente el Mar Muerto, Jordania (Tabla complementaria S4) proporciona una idea de la ruta migratoria de regreso a África.
Referencias
^ abc Fu Q, Posth C, Hajdinjak M, Petr M, Mallick S, Fernandes D, et al. (junio de 2016). "La historia genética de la Edad de Hielo en Europa". Nature . 534 (7606): 200–5. Bibcode :2016Natur.534..200F. doi :10.1038/nature17993. hdl :10211.3/198594. PMC 4943878 . PMID 27135931.
^ abcdef Haak y otros 2015.
^ ab Allentoft ME, Sikora M, Sjögren KG, Rasmussen S, Rasmussen M, Stenderup J, et al. (junio de 2015). "Genómica de poblaciones de Eurasia de la Edad del Bronce". Naturaleza . 522 (7555): 167–72. Código Bib :2015Natur.522..167A. doi : 10.1038/naturaleza14507. PMID 26062507. S2CID 4399103.
^ ab Mathieson I, Lazaridis I, Rohland N, Mallick S, Patterson N, Roodenberg SA, et al. (2015). "Ocho mil años de selección natural en Europa". bioRxiv : 016477. doi :10.1101/016477. S2CID 7866359.
^ Cassidy LM, Martiniano R, Murphy EM, Teasdale MD, Mallory J, Hartwell B, Bradley DG (enero de 2016). "Migración neolítica y de la Edad del Bronce a Irlanda y establecimiento del genoma atlántico insular". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 113 (2): 368–73. Bibcode :2016PNAS..113..368C. doi : 10.1073/pnas.1518445113 . PMC 4720318 . PMID 26712024.
^ Martiniano R, Cassidy LM, Ó'Maoldúin R, McLaughlin R, Silva NM, Manco L, et al. (julio de 2017). "La genómica poblacional de la transición arqueológica en el oeste de Iberia: investigación de la subestructura antigua utilizando métodos de imputación y basados en haplotipos". PLOS Genetics . 13 (7): e1006852. doi : 10.1371/journal.pgen.1006852 . PMC 5531429 . PMID 28749934.
^ abc Karafet TM, Mendez FL, Meilerman MB, Underhill PA, Zegura SL, Hammer MF (mayo de 2008). "Nuevos polimorfismos binarios remodelan y aumentan la resolución del árbol de haplogrupos del cromosoma Y humano". Genome Research . 18 (5): 830–8. doi :10.1101/gr.7172008. PMC 2336805 . PMID 18385274.
^ Mathieson 2018, Tabla suplementaria 1, fila 467.
^ Mathieson 2018, Tabla suplementaria 1, filas 251-272.
^Por Jones 2017.
^ Mathieson 2018, Tabla suplementaria 1, filas 205-245.
^ Mathieson 2018, Tabla suplementaria 1.
^ Mathieson 2018, págs. 2-3.
^ González-Fortes et al. 2017, págs. 8–9.
^ González-Fortes et al. 2017, pág. 4, Tabla 1, OC1_Meso.
^ Sánchez-Quinto et al. 2019, conjunto de datos ampliado 1.3, OC1.
^ Mathieson 2018, Tabla suplementaria 1, fila 298, I4666.
^ Mathieson 2018, Tabla suplementaria 1, fila 153, I0124.
^ Mathieson 2018, Tabla suplementaria 1, fila 375, I0122.
^ Haak et al. 2015, Información complementaria, págs. 26-27.
^ Haak y col. 2015, Tabla de datos ampliada 2, I0410.
^ Haak et al. 2015, Información complementaria, págs. 44-45.
^ Mathieson 2018, Tabla suplementaria 1, I0410.
^ Mathieson 2018, Tabla suplementaria 1, fila 73, I2430.
^ Marcus et al. 2020, Datos suplementarios 1, Tabla maestra, Fila 25, MA89.
^ Mathieson 2018, Tabla suplementaria 1, fila 128, I0559.
^ de Barros Damgaard P, Martiniano R, Kamm J, Moreno-Mayar JV, Kroonen G, Peyrot M, et al. (junio de 2018). "Los primeros pastores de caballos y el impacto de las expansiones esteparias de la Edad del Bronce Temprano en Asia". Science . 360 (6396): eaar7711. doi :10.1126/science.aar7711. PMC 6748862 . PMID 29743352.
^ Haak y otros. 2015, pág. 5.
^ Karafet TM, Mendez FL, Sudoyo H, Lansing JS, Hammer MF (marzo de 2015). "Mejora de la resolución filogenética y rápida diversificación del haplogrupo K-M526 del cromosoma Y en el sudeste asiático". Revista Europea de Genética Humana . 23 (3): 369–73. doi :10.1038/ejhg.2014.106. PMC 4326703 . PMID 24896152.
^ Sikora, Martín; Pitulko, Vladimir V.; Sousa, Vítor C.; Allentoft, Morten E.; Vinner, Lasse; Rasmussen, Simón; Margaryan, Ashot; de Barros Damgaard, Peter; de la Fuente, Constanza; Renaud, Gabriel; Yang, Melinda A.; Fu, Qiaomei; Dupanloup, Isabelle; Giampoudakis, Konstantinos; Nogués-Bravo, David (junio de 2019). "La historia de la población del noreste de Siberia desde el Pleistoceno". Naturaleza . 570 (7760): 182–188. Código Bib :2019Natur.570..182S. doi :10.1038/s41586-019-1279-z. hdl : 1887/3198847 . Revista de Biología Molecular y Genética .
^ Lacan M, Keyser C, Ricaut FX, Brucato N, Duranthon F, Guilaine J, et al. (junio de 2011). "El ADN antiguo revela la difusión masculina a través de la ruta mediterránea neolítica". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 108 (24): 9788–91. Bibcode :2011PNAS..108.9788L. doi : 10.1073/pnas.1100723108 . PMC 3116412 . PMID 21628562.
^ Haak W, Balanovsky O, Sanchez JJ, Koshel S, Zaporozhchenko V, Adler CJ, et al. (noviembre de 2010). Penny D (ed.). "El ADN antiguo de los agricultores neolíticos europeos revela sus afinidades con el Cercano Oriente". PLOS Biology . 8 (11): e1000536. doi : 10.1371/journal.pbio.1000536 . PMC 2976717 . PMID 21085689.
^ Olalde I, Brace S, Allentoft ME, Armit I, Kristiansen K, Booth T, et al. (marzo de 2018). "El fenómeno Beaker y la transformación genómica del noroeste de Europa". Nature . 555 (7695): 190–196. Bibcode :2018Natur.555..190O. doi :10.1038/nature25738. PMC 5973796 . PMID 29466337.
^ abc Cinnioğlu C, King R, Kivisild T, Kalfoğlu E, Atasoy S, Cavalleri GL, et al. (enero de 2004). "Excavando los estratos de haplotipos del cromosoma Y en Anatolia". Genética humana . 114 (2): 127–48. doi :10.1007/s00439-003-1031-4. PMID 14586639. S2CID 10763736.
^ ab "ISOGG 2016 Y-ADN Haplogrupo R". isogg.org .
^ abc "ISOGG 2017 Haplogrupo R de ADN-Y". isogg.org .
^ YFull YTree v10.00.00
^ Árbol filogenético del haplogrupo R-V1636 en 23mofang
^ Haplogrupo R en el haploárbol de ADN-Y en FTDNA
^ "R-V1636 YTree". www.yfull.com .
^ abcdefghijklm Myres NM, Rootsi S, Lin AA, Järve M, King RJ, Kutuev I, et al. (Enero de 2011). "Un importante efecto fundador de la era del Holoceno del haplogrupo R1b del cromosoma Y en Europa central y occidental". Revista europea de genética humana . 19 (1): 95-101. doi :10.1038/ejhg.2010.146. PMC 3039512 . PMID 20736979.
^ Grugni V, Battaglia V, Hooshiar Kashani B, Parolo S, Al-Zahery N, Achilli A, et al. (18 de julio de 2012). "Antiguos acontecimientos migratorios en Oriente Medio: nuevas pistas a partir de la variación del cromosoma Y de los iraníes modernos". PLOS ONE . 7 (7): e41252. Bibcode :2012PLoSO...741252G. doi : 10.1371/journal.pone.0041252 . PMC 3399854 . PMID 22815981.
^ Adams SM, King TE, Bosch E, Jobling MA (mayo de 2006). "El caso del SNP no confiable: mutación inversa recurrente del marcador del cromosoma Y P25 a través de la conversión génica". Forensic Science International . 159 (1): 14–20. doi :10.1016/j.forsciint.2005.06.003. hdl : 2381/443 . PMID 16026953.
^ abc Cruciani F, Trombetta B, Sellitto D, Massaia A, Destro-Bisol G, Watson E, et al. (Julio de 2010). "Haplogrupo R-V88 del cromosoma Y humano: un registro genético paterno de las conexiones transaharianas del Holoceno medio temprano y la propagación de las lenguas chadic". Revista europea de genética humana . 18 (7): 800–7. doi :10.1038/ejhg.2009.231. PMC 2987365 . PMID 20051990.
^ Varzari A (2006). Historia de la población de los Cárpatos Dniéster: evidencia de la inserción de Alu y polimorfismos del cromosoma Y (Tesis). Ludwig-Maximilians-Universität München. doi :10.5282/edoc.5868.
^ ab Malyarchuk B, Derenko M, Denisova G, Maksimov A, Wozniak M, Grzybowski T, et al. (agosto de 2011). "Antiguos vínculos entre siberianos y nativos americanos revelados mediante la subtipificación del haplogrupo Q1a del cromosoma Y". Journal of Human Genetics . 56 (8): 583–8. doi : 10.1038/jhg.2011.64 . PMID 21677663. S2CID 12015336.
^ Dulik MC, Zhadanov SI, Osipova LP, Askapuli A, Gau L, Gokcumen O, et al. (febrero de 2012). "La variación del ADN mitocondrial y del cromosoma Y proporciona evidencia de una ascendencia común reciente entre los nativos americanos y los indígenas altaianos". American Journal of Human Genetics . 90 (2): 229–46. doi :10.1016/j.ajhg.2011.12.014. PMC 3276666 . PMID 22281367.
^ Behar DM, Yunusbayev B, Metspalu M, Metspalu E, Rosset S, Parik J, et al. (julio de 2010). "La estructura de todo el genoma del pueblo judío". Nature . 466 (7303): 238–42. Bibcode :2010Natur.466..238B. doi :10.1038/nature09103. PMID 20531471. S2CID 4307824.
^ Sengupta S, Zhivotovsky LA, King R, Mehdi SQ, Edmonds CA, Chow CE, et al. (febrero de 2006). "La polaridad y la temporalidad de las distribuciones de alta resolución del cromosoma Y en la India identifican expansiones tanto indígenas como exógenas y revelan una influencia genética menor de los pastores de Asia central". American Journal of Human Genetics . 78 (2): 202–21. doi :10.1086/499411. PMC 1380230 . PMID 16400607.
^ ab Di Cristofaro J, Pennarun E, Mazières S, Myres NM, Lin AA, Temori SA, et al. (2013). "Afghan Hindu Kush: donde convergen los flujos de genes del subcontinente euroasiático". MÁS UNO . 8 (10): e76748. Código Bib : 2013PLoSO...876748D. doi : 10.1371/journal.pone.0076748 . PMC 3799995 . PMID 24204668.
^ Lippold S, Xu H, Ko A, Li M, Renaud G, Butthof A, et al. (2014). "Historias demográficas maternas y paternas humanas: perspectivas a partir de secuencias de alta resolución del cromosoma Y y del ADNmt". Genética investigativa . 5 : 13. doi : 10.1186/2041-2223-5-13 . PMC 4174254 . PMID 25254093.
^ Karafet TM, Osipova LP, Savina OV, Hallmark B, Hammer MF (noviembre de 2018). "La diversidad genética siberiana revela orígenes complejos de las poblaciones de habla samoyeda". American Journal of Human Biology . 30 (6): e23194. doi :10.1002/ajhb.23194. PMID 30408262. S2CID 53238849.
^ Ashirbekov EE, Botbaev DM, Belkozhaev AM, Abayldaev AO, Neupokoeva AS, Mukhataev JE, et al. (2017). "Распределение гаплогрупп И-хромосомы казахов Южно-Казахстанской, Жамбылской и Алматинской областей" [Distribución de los haplogrupos del cromosoma Y del kazajo del sur Regiones de Kazajstán, Zhambyl y Almaty] (PDF) . Informes de la Academia Nacional de Ciencias de la República de Kazajstán (en ruso). 6 (316): 85–95. Archivado desde el original (PDF) el 23 de septiembre de 2021. Consultado el 20 de noviembre de 2021 .
^ Shuhu LI, Yilihamu NI, Bake RA, Bupatima AB, Matyusup DO (2018). "Un estudio de la diversidad genética de tres poblaciones aisladas en Xinjiang utilizando Y-SNP". Acta Antropológica Sínica . 37 (1): 146–56. doi : 10.16359/j.cnki.cn11-1963/q.2017.0067.
^ R1b1a1b (R-M269) era anteriormente R1b1a1a2. De 2003 a 2005, lo que ahora es R1b1a1b se denominó R1b3. De 2005 a 2008, fue R1b1c. De 2008 a 2011, fue R1b1b2. De 2011 a 2018, fue R1b1a1a2.
^ ab Balaresque P, Bowden GR, Adams SM, Leung HY, King TE, Rosser ZH, et al. (enero de 2010). Penny D (ed.). "Un origen predominantemente neolítico para los linajes paternos europeos". PLOS Biology . 8 (1): e1000285. doi : 10.1371/journal.pbio.1000285 . PMC 2799514 . PMID 20087410.
^ Arredi B, Poloni ES, Tyler-Smith C (2007). "El poblamiento de Europa". En Crawford MH (ed.). Genética antropológica: teoría, métodos y aplicaciones . Cambridge, Reino Unido: Cambridge University Press. p. 394. ISBN978-0-521-54697-3.
^ Cruciani F, Trombetta B, Antonelli C, Pascone R, Valesini G, Scalzi V, et al. (junio de 2011). "Fuerte diferenciación intra e intercontinental revelada por los SNP del cromosoma Y M269, U106 y U152". Forensic Science International. Genética . 5 (3): e49-52. doi :10.1016/j.fsigen.2010.07.006. hdl :11573/226727. PMID 20732840.
^ Grugni, Viola; Raveane, Alessandro; Mattioli, Francesca; Battaglia, Vincenza; Sala, Cinzia; Toniolo, Daniela; Ferretti, Luca; Gardella, Rita; Aquiles, Alejandro; Olivieri, Anna; Torroní, Antonio; Passarino, Giuseppe; Semino, Ornella (febrero de 2018). "Reconstrucción de la historia genética de los italianos: nuevos conocimientos desde una perspectiva masculina (cromosoma Y)". Anales de biología humana . 45 (1): 44–56. doi :10.1080/03014460.2017.1409801. ISSN 1464-5033. PMID 29382284. S2CID 43501209.
^ ab Underhill PA, Shen P, Lin AA, Jin L, Passarino G, Yang WH, et al. (noviembre de 2000). "Variación de la secuencia del cromosoma Y y la historia de las poblaciones humanas". Nature Genetics . 26 (3): 358–361. doi :10.1038/81685. PMID 11062480. S2CID 12893406.
^ Lobov AS (2009). Estructura del acervo genético de las subpoblaciones bashkires (PDF) (tesis doctoral) (en ruso). Instituto de Bioquímica y Genética del Centro Científico de Ufa de la Academia Rusa de Ciencias. Archivado desde el original (PDF) el 16 de agosto de 2011.
^ Ordóñez AC, Fregel R, Trujillo-Mederos A, Hervella M, de-la-Rúa C, Arnay-de-la-Rosa M (2017). "Estudios genéticos sobre la población prehispánica enterrada en la cueva de Punta Azul (El Hierro, Canarias)". Revista de Ciencias Arqueológicas . 78 : 20–28. Código Bib : 2017JArSc..78...20O. doi :10.1016/j.jas.2016.11.004.
^ Robino C, Crobu F, Di Gaetano C, Bekada A, Benhamamouch S, Cerutti N, et al. (mayo de 2008). "Análisis de haplogrupos de SNP del cromosoma Y y haplotipos de STR en una muestra de población argelina". Revista Internacional de Medicina Legal . 122 (3): 251–5. doi :10.1007/s00414-007-0203-5. PMID 17909833. S2CID 11556974.
^ abc Wood ET, Stover DA, Ehret C, Destro-Bisol G, Spedini G, McLeod H, et al. (julio de 2005). "Patrones contrastantes de variación del cromosoma Y y del ADNmt en África: evidencia de procesos demográficos con sesgo sexual". Revista Europea de Genética Humana . 13 (7): 867–76. doi : 10.1038/sj.ejhg.5201408 . PMID 15856073. S2CID 20279122.
^ Yepiskoposian L, Khudoyan A, Harutyunian A (2006). "Prueba genética de la hipótesis de reemplazo del lenguaje en el suroeste de Asia". Irán y el Cáucaso . 10 (2): 191–208. doi :10.1163/157338406780345899. JSTOR 4030922. S2CID 162345193.
^ Vadimovna TN (febrero de 2015). Изменчивость Митохондриальной ДНК и Y-Хромосомы в Популяциях Волго-Уральского Региона [ Variación del ADN mitocondrial y el cromosoma Y en la población de la región Volga-Ural ] (Tesis de Ciencias Biológicas) (en ruso). Archivado desde el original (PDF) el 2 de abril de 2017.
^ Herrera KJ, Lowery RK, Hadden L, Calderon S, Chiou C, Yepiskoposyan L, et al. (marzo de 2012). "Las señales patrilineales neolíticas indican que la meseta armenia fue repoblada por agricultores". Revista Europea de Genética Humana . 20 (3): 313–20. doi :10.1038/ejhg.2011.192. PMC 3286660 . PMID 22085901.
^ Vanek D, Saskova L, Koch H (junio de 2009). "Análisis de parentesco y cromosoma Y de restos humanos del siglo VII: nuevo procedimiento de extracción y tipificación de ADN para material antiguo". Revista Médica Croata . 3. 50 (3): 286–95. doi :10.3325/cmj.2009.50.286. PMC 2702742 . PMID 19480023.
^ D'Atanasio E, Trombetta B, Bonito M, Finocchio A, Di Vito G, Seghizzi M, et al. (febrero de 2018). "El poblamiento del último Sahara Verde revelado por la resecuenciación de alta cobertura de linajes patriarcales transsaharianos". Genome Biology . 19 (1): 20. doi : 10.1186/s13059-018-1393-5 . PMC 5809971 . PMID 29433568.
^ Marcus JH, Posth C, Ringbauer H, Lai L, Skeates R, Sidore C, et al. (febrero de 2020). "Historia genética desde el Neolítico Medio hasta la actualidad en la isla mediterránea de Cerdeña". Comunicaciones de la naturaleza . 11 (1): 939. Código bibliográfico : 2020NatCo..11..939M. doi :10.1038/s41467-020-14523-6. PMC 7039977 . PMID 32094358.
^ ab Grugni, Viola; et al. (2019). "Análisis del cromosoma Y y del apellido para reconstruir estructuras de poblaciones pasadas: la población sarda como caso de prueba". Revista internacional de ciencias moleculares . 20 (5763): 5763. doi : 10.3390/ijms20225763 . PMC 6888588 . PMID 31744094.
^ Allentoft, ME (2024). "Genómica de poblaciones de Eurasia occidental postglacial". Nature . 625 (7994): Información suplementaria, p.48. Bibcode :2024Natur.625..301A. doi :10.1038/s41586-023-06865-0. PMC 10781627 . PMID 38200295.
^ Haber, Marc; Mezzavilla, Massimo; Bergstrom, Anders; Prado-Martínez, Javier; Hallast, Pille; Saif-Ali, Riad; Al-Habori, Molham; Dedoussis, George; Zeggini, Eleftheria; Blue-Smith, Jason; Wells, R. Spencer; Xue, Yalí; Zalloua, Pierre A.; Tyler-Smith, Chris (diciembre de 2016). "La diversidad genética de Chad revela una historia africana marcada por múltiples migraciones euroasiáticas del Holoceno" (PDF) . La Revista Estadounidense de Genética Humana . 99 (6): 1316-1324. doi :10.1016/j.ajhg.2016.10.012. PMC 5142112 . Número de modelo: PMID 27889059. Número de modelo: S2CID 38169172.
^ Kulichova, Iva; et al. (2017). "Diversificación interna de haplogrupos no subsaharianos en poblaciones sahelianas y la expansión del pastoralismo más allá del Sahara". Revista estadounidense de antropología biológica . 164 (2): 424–434. doi :10.1002/ajpa.23285. PMID 28736914.
^ Vicente, Mario; et al. (2019). "Historia poblacional y adaptación genética de los nómadas Fulani: inferencias a partir de datos de todo el genoma y el rasgo de persistencia de la lactasa". BMC Genomics . 20 (1): 915. doi : 10.1186/s12864-019-6296-7 . PMC 6888939 . PMID 31791255.
^ Simões, LG; et al. (2023). "Neolítico del noroeste de África iniciado por migrantes de Iberia y el Levante". Nature . 618 (7965): 550–556. Bibcode :2023Natur.618..550S. doi :10.1038/s41586-023-06166-6. PMC 10266975 . PMID 37286608.
^ Fregel, Rosa; Méndez, Fernando L.; Bokbot, Youssef; Martín-Socas, Dimas; Camalich-Massieu, María D.; Santana, Jonatán; Morales, Jacob; Ávila-Arcos, María C.; Underhill, Peter A.; Shapiro, Beth; Wojcik, Genevieve; Rasmussen, Morten; Soares, André ER; Kapp, Josué; Sockell, Alexandra (26 de junio de 2018). "Los genomas antiguos del norte de África evidencian migraciones prehistóricas al Magreb tanto desde el Levante como desde Europa". Actas de la Academia Nacional de Ciencias . 115 (26): 6774–6779. Código Bib : 2018PNAS..115.6774F. doi : 10.1073/pnas.1800851115 . ISSN 0027-8424. Número de modelo : PMID 29895688 .
^ Flores C, Maca-Meyer N, Larruga JM, Cabrera VM, Karadsheh N, Gonzalez AM (2005). "Aislamientos en un corredor de migraciones: un análisis de alta resolución de la variación del cromosoma Y en Jordania". Journal of Human Genetics . 50 (9): 435–441. doi : 10.1007/s10038-005-0274-4 . PMID 16142507. S2CID 6490283.
^ Hassan HY, Underhill PA, Cavalli-Sforza LL, Ibrahim ME (noviembre de 2008). "Variación del cromosoma Y entre los sudaneses: flujo genético restringido, concordancia con el idioma, la geografía y la historia". American Journal of Physical Anthropology . 137 (3): 316–323. doi :10.1002/ajpa.20876. PMID 18618658. 13/32
^ Shriner, Daniel; Rotimi, Charles N. (diciembre de 2018). "Historia genética de Chad". Revista estadounidense de antropología física . 167 (4): 804–812. doi :10.1002/ajpa.23711. ISSN 0002-9483. PMC 6240361 . PMID 30259956.
^ Cruciani F, Santolamazza P, Shen P, Macaulay V, Moral P, Olckers A, et al. (mayo de 2002). "Una migración de regreso desde Asia al África subsahariana está respaldada por un análisis de alta resolución de los haplotipos del cromosoma Y humano". American Journal of Human Genetics . 70 (5): 1197–214. doi :10.1086/340257. PMC 447595 . PMID 11910562. , págs. 13-14
^ Contu D, Morelli L, Santoni F, Foster JW, Francalacci P, Cucca F (enero de 2008). "Evidencia basada en el cromosoma Y del origen preneolítico de la población sarda, genéticamente homogénea pero diversa: inferencia para exploraciones de asociación". PLOS ONE . 3 (1): e1430. Bibcode :2008PLoSO...3.1430C. doi : 10.1371/journal.pone.0001430 . PMC 2174525 . PMID 18183308.
^ Zalloua PA, Xue Y, Khalife J, Makhoul N, Debiane L, Platt DE, et al. (Abril de 2008). "La diversidad del cromosoma Y en el Líbano está estructurada por acontecimientos históricos recientes". Revista Estadounidense de Genética Humana . 82 (4): 873–882. doi :10.1016/j.ajhg.2008.01.020. PMC 2427286 . PMID 18374297.
^ Zhong H, Shi H, Qi XB, Duan ZY, Tan PP, Jin L, et al. (enero de 2011). "Una investigación ampliada del cromosoma Y sugiere migraciones postglaciales de humanos modernos hacia el este de Asia a través de la ruta del norte". Biología molecular y evolución . 28 (1): 717–27. doi : 10.1093/molbev/msq247 . PMID 20837606.
^ Rowold DJ, Perez Benedico D, Garcia-Bertrand R, Chennakrishnaiah S, Alfonso-Sanchez MA, Gayden T, Herrera RJ (marzo de 2016). "Ladakh, India: la tierra de los altos pasos y la heterogeneidad genética revela una confluencia de migraciones". Revista Europea de Genética Humana . 24 (3): 442–9. doi :10.1038/ejhg.2015.80. PMC 4755386 . PMID 25966630.
^ "Sobre R1a y R1b de la historia épica de los Urales. Artem Lukichev (c)". 5 de agosto de 2014. Archivado desde el original el 15 de diciembre de 2021 – vía www.youtube.com.
^ Coble MD, Loreille OM, Wadhams MJ, Edson SM, Maynard K, Meyer CE, et al. (2009). "Misterio resuelto: la identificación de los dos niños Romanov desaparecidos mediante análisis de ADN". PLOS ONE . 4 (3): e4838. Bibcode :2009PLoSO...4.4838C. doi : 10.1371/journal.pone.0004838 . PMC 2652717 . PMID 19277206.
^ "¿Odkud přišli Přemyslovci? Analýza DNA byla pro vědce velkým překvapením". dotyk.cz (en checo) . Consultado el 13 de mayo de 2023 .
^ Larmuseau, Maarten HD; Delorme, Philippe; Germain, Patrick; Vanderheyden, Nancy (9 de octubre de 2013). "La genealogía genética revela el verdadero haplogrupo Y de la Casa de Borbón, lo que contradice la reciente identificación de los presuntos restos de dos reyes franceses". Eur J Hum Genet . 22 (5): 681–687. doi :10.1038/ejhg.2013.211. PMC 3992573 . PMID 24105374.
^ Gad, Yehia (2020). "Linajes paternos y maternos en la familia del rey Tutankamón". Guardián del Antiguo Egipto: Ensayos en honor a Zahi Hawass. Instituto Checo de Egiptología. pp. 497–518. ISBN978-80-7308-979-5.
^ Gad, Yehia (2020). "Información obtenida a partir del análisis del ADN antiguo de momias humanas egipcias: pistas sobre enfermedades y parentesco". Genética molecular humana . 30 (R1): R24–R28. doi : 10.1093/hmg/ddaa223 . ISSN 0964-6906. PMID 33059357.
^ Maan AA, Eales J, Akbarov A, Rowland J, Xu X, Jobling MA, et al. (noviembre de 2017). "El cromosoma Y: ¿un modelo para la salud de los hombres?". Revista europea de genética humana . 25 (11): 1181–1188. doi :10.1038/ejhg.2017.128. PMC 5643963 . PMID 28853720.
^ Timmers P, Wilson JF (julio de 2022). "Efecto limitado de la variación del cromosoma Y en la enfermedad coronaria y la mortalidad en el Biobanco del Reino Unido: informe breve". Arteriosclerosis, trombosis y biología vascular . 42 (9): 1198–1206. doi :10.1161/ATVBAHA.122.317664. PMC 9394501 . PMID 35861954.
Bibliografía
Jones ER, Zarina G, Moiseyev V, Lightfoot E, Nigst PR, Manica A, et al. (febrero de 2017). "La transición neolítica en el Báltico no fue impulsada por la mezcla con los primeros agricultores europeos". Current Biology . 27 (4): 576–582. Bibcode :2017CBio...27..576J. doi :10.1016/j.cub.2016.12.060. PMC 5321670 . PMID 28162894.
González-Fortes G, Jones ER, Lightfoot E, Bonsall C, Lazar C, Grandal-d'Anglade A, et al. (junio de 2017). "Evidencia paleogenómica de mezcla multigeneracional entre agricultores neolíticos y cazadores-recolectores mesolíticos en la cuenca baja del Danubio". Current Biology . 27 (12): 1801–1810.e10. Bibcode :2017CBio...27E1801G. doi :10.1016/j.cub.2017.05.023. PMC 5483232 . PMID 28552360.
Haak W, Lazaridis I, Patterson N, Rohland N, Mallick S, Llamas B, et al. (junio de 2015). "La migración masiva desde la estepa fue una fuente de lenguas indoeuropeas en Europa". Nature . 522 (7555): 207–211. arXiv : 1502.02783 . Bibcode :2015Natur.522..207H. doi :10.1038/nature14317. PMC 5048219 . PMID 25731166.
Hollard C, Zvénigorosky V, Kovalev A, Kiryushin Y, Tishkin A, Lazaretov I, et al. (septiembre de 2018). "Nueva evidencia genética de afinidades y discontinuidades entre poblaciones siberianas de la Edad del Bronce". Revista estadounidense de antropología física . 167 (1): 97–107. doi :10.1002/ajpa.23607. PMID 29900529. S2CID 205337212.
Lipson M, Szécsényi-Nagy A, Mallick S, Pósa A, Stégmár B, Keerl V, et al. (noviembre de 2017). "Los transectos paleogenómicos paralelos revelan una historia genética compleja de los primeros agricultores europeos". Naturaleza . 551 (7680): 368–372. Código Bib :2017Natur.551..368L. doi : 10.1038/naturaleza24476. PMC 5973800 . PMID 29144465.
Marcus JH, Posth C, Ringbauer H, Lai L, Skeates R, Sidore C, et al. (febrero de 2020). "Historia genética desde el Neolítico Medio hasta la actualidad en la isla mediterránea de Cerdeña". Comunicaciones de la naturaleza . 11 (1): 939. Código bibliográfico : 2020NatCo..11..939M. doi :10.1038/s41467-020-14523-6. PMC 7039977 . PMID 32094358.
Mathieson I, Alpaslan-Roodenberg S, Posth C, Szécsényi-Nagy A, Rohland N, Mallick S, et al. (Marzo de 2018). "La historia genómica del sureste de Europa". Naturaleza . 555 (7695): 197–203. Código Bib :2018Natur.555..197M. doi : 10.1038/naturaleza25778. PMC 6091220 . PMID 29466330.
Narasimhan VM, Patterson N, Moorjani P, Rohland N, Bernardos R, Mallick S, et al. (septiembre de 2019). "La formación de poblaciones humanas en Asia central y meridional". Ciencia . 365 (6457): comer7487. doi : 10.1126/science.aat7487. PMC 6822619 . PMID 31488661.
Olalde I, Brace S, Allentoft ME, Armit I, Kristiansen K, Booth T, et al. (marzo de 2018). "El fenómeno Beaker y la transformación genómica del noroeste de Europa". Nature . 555 (7695): 190–196. Bibcode :2018Natur.555..190O. doi :10.1038/nature25738. PMC 5973796 . PMID 29466337.
Haak, Wolfgang; Lazaridis, José; Patterson, Nick; Rohland, Nadin; Mallick, Swapan; Llamas, Bastien; Brandt, Guido; Nordenfelt, Susanne; et al. (2015). "La migración masiva desde la estepa es una fuente de lenguas indoeuropeas en Europa". Naturaleza . 522 (7555): 207–211. arXiv : 1502.02783 . Código Bib :2015Natur.522..207H. bioRxiv 10.1101/013433 . doi :10.1038/NATURALEZA14317. PMC 5048219 . PMID 25731166.
Sánchez-Quinto F, Malmström H, Fraser M, Girdland-Flink L, Svensson EM, Simões LG, et al. (mayo de 2019). "Las tumbas megalíticas en el Neolítico occidental y septentrional de Europa estaban vinculadas a una sociedad afín". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 116 (19): 9469–9474. Bibcode :2019PNAS..116.9469S. doi : 10.1073/pnas.1818037116 . PMC 6511028 . PMID 30988179.
Fu Q, Posth C, Hajdinjak M, Petr M, Mallick S, Fernandes D, et al. (junio de 2016). "La historia genética de la Edad de Hielo en Europa". Nature . 534 (7606): 200–5. Bibcode :2016Natur.534..200F. doi :10.1038/nature17993. PMC 4943878 . PMID 27135931.