SNARE (proteína)
[3] Las SNARE más estudiadas son las que median el acoplamiento de las vesículas sinápticas con la membrana presináptica en las neuronas.
[5] Varias proteínas SNARE se encuentran tanto en las vesículas como en las membranas diana, por lo que un esquema de clasificación más reciente toma en cuenta las características estructurales de las SNARE, dividiéndolas en R-SNARE y Q-SNARE.
Las Q-SNARE se pueden clasificar a su vez como Qa, Qb o Qc, dependiendo de su ubicación en el complejo.
Existen excepciones como la brevina, sin dominios adicionales en el extremo amino, y la SNAP-25, que carece de dominio transmembranal pero que se une a la membrana por palmitoilación en la región conectora entre sus dos motivos SNARE.
La adaptación de las SNARE a su membrana diana se logra mediante diferencias en los aminoácidos que rodean el extremo carboxilo o en la longitud del dominio transmembrana.
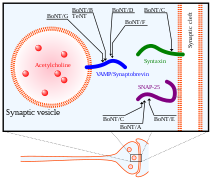
En las vesículas sinápticas, estos complejos «trans» son extremadamente estables y se componen de tres SNARE: sintaxina y SNAP-25, ancladas a la membrana celular y sinaptobrevina —también conocida como proteína de membrana asociada a vesículas o VAMP— asociada a la membrana de la vesícula.
Estas capas presentan carácter hidrofóbico, excepto la capa 0, en la posición central del complejo, compuesta por una arginina (R) —procedente de la sinaptobrevina— que interactúa con tres glutaminas (Q) aportadas por la sintaxina, Sn1 y Sn2 respectivamente.
Las proteínas SNARE generan energía a través interacciones proteína-lípidos y proteína-proteína, que actúan como una fuerza impulsora para la fusión de membrana.
[6] El proceso de ensamblaje está limitado por el dominio SNARE de la sintaxina, que habitualmente se encuentra unida a la proteína Munc18 en una conformación que le imposibilita interactuar con otras proteínas SNARE.
Existen varias hipótesis sobre estos procesos, aunque su naturaleza exacta y el papel que desempeñan las proteínas SNARE es objeto de debate.
Se sospecha que el dominio transmembrana de las SNARE contribuye al acercamiento, quizá produciendo un abultamiento local de la membrana, porque el tallo no se forma cuando este dominio es reemplazado por lípidos.
El NSF no se une directamente a las proteínas SNARE, sino que actúa por medio de su cofactor alpha-SNAP.
[6][19] Este proceso permite la recaptación de la sinaptobrevina para su uso posterior en las vesículas, mientras que las otras proteínas SNARE permanecen asociadas a la membrana celular.
La proteína Q-SNARE asociada al sinaptosoma (SNAP-25) consta de dos dominios alpha-helicoidales conectados por un segmento sin estructura secundaria definida que contiene cuatro cisteínas.
Sin embargo, los experimentos con sintaxina inactivada mediante modificación genética no han hallado una disminución en la unión de SNAP-25 a la membrana, lo que sugiere que el medio de conexión directo existe.
El proceso inverso se lleva a cabo mediante otra enzima llamada palmitoil proteína tioesterasa o PPT.
[26] Se cree que la proteína Munc-18 puede estabilizar tanto la conformación cerrada como el complejo SNARE-trans.
[29] Muchas neurotoxinas interfieren directamente con el complejo SNARE; las toxinas botulínica y tetánica, por ejemplo, impiden el reciclaje de vesículas y provocan la pérdida de control muscular, espasmos, parálisis e incluso la muerte.
[31] Hay ocho isotipos conocidos de BoNT, BoNT/A-H, que escinden las proteínas SNARE en diferentes sitios.
Aunque los efectos de la neurotoxina botulínica pueden ser letales, también se ha utilizado como agente terapéutico en algunos tratamientos médicos y cosméticos.
Cuando las vesículas entran en contacto físico con la membrana, la sintaxina 1A, previamente inhibida por Munc-18, queda libre para unirse a las proteínas v-SNARE.
Los canales de calcio dependientes del voltaje se concentran en torno a las zonas activas.
[46] Las proteínas SNARE tienen un papel importante en la fusión de vesículas en la formación del fagoforo.
[48] Las SNAREs también son importantes en la mediación de la fusión del autofagosoma y el lisosoma.