Spermatophyta
Debido a que en las espermatófitas el grano de polen produce un tubo (haustorial o polínico) para llegar al óvulo y que ocurra la fecundación, este grupo también es llamado embriofitas sifonógamas, del griego embrios: «embrión»; fiton: «planta»; xifos: «tubo»; gamos: «unión sexual».
Hoy en día, las espermatófitas conforman el linaje más extenso de plantas vasculares, con unas 270.000 especies vivientes.
[1] Un solo subclado es el mayor responsable de esa diversidad: las angiospermas o plantas con flores periantadas.
Otros subclados, normalmente agrupados como gimnospermas, son las cícadas, los ginkgos, las coníferas y los gnetales.
Este grupo ha sido denominado Phanerogama (Bartling 1830), Phanerogamae (Brongniart 1843, Eichler 1883), Spermatophyta (Willkomm 1854, Goebel 1882, Britton & A. Brown 1896), Anthophyta (Braun in Ascherson 1864, Wettstein 1924-1935), Siphonogamae (Engler 1886), Embryophyta siphonogama, (Engler 1892-1924), división Magnoliophyta (Takhtajan 1964) y Spermatophytina (Cavalier-Smith 1998, Ruggiero et al.
Recordemos que la diferenciación entre un tallo principal y ramas laterales ya había evolucionado en el linaje de las eufilofitas.
Estos troncos estaban conectados a enormes sistemas de ramas que portaban muchas hojas pequeñas.
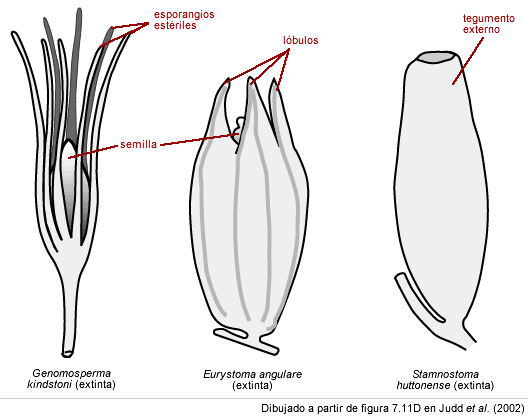
Análisis cuidadosos (por ejemplo el de Serbet y Rothwell 1992) han revelado que las primeras semillas estaban situadas en "cúpulas" ("cupules" en inglés) y cada semilla era cubierta por una excrecencia de la pared del esporangio que formaba una cámara especializada para recibir el polen, o "cámara polínica".
[2] Así pues, la filogenia más actualizada (2014) de las plantas vivientes con semilla presenta los siguientes subgrupos:[3]
Esta distinción sugirió a algunos investigadores que las plantas con semilla en realidad se originaron dos veces.
Este escenario implica que la semilla misma fue originada dos veces, cada una correspondiéndose a un tipo de simetría diferente.
Sin embargo, en los análisis de filogenia que incluyeron los linajes vivientes junto con los representantes fósiles, en general aprueban el árbol filogenético que se muestra en la figura siguiente (ver por ejemplo Crane 1985, Doyle y Donoghue 1986, Nixon et al.
Por lo tanto Cycas estaría reteniendo algunos caracteres presumiblemente ancestrales, como los que han sido encontrados en parientes fósiles como Taeniopteris, a saber: el tener muchos óvulos nacidos en los márgenes de carpelos (carpelos definidos como hojas fértiles portadoras de óvulos) con mofología de tipo foliar, en lugar de tener dos óvulos por carpelo peltado que los sostiene apuntando hacia el eje del esporofilo (en su "cara adaxial"), que es el carácter que se encuentra en la otra línea.
También en Cycas las hojas fértiles portadoras de óvulos no se encuentran agrupadas en estróbilos, como sí lo están en la otra línea.
Los ginkgos son bien conocidos en el registro fósil, donde se observa una gran diversidad en la morfología de las hojas.
También al igual que en las cícadas, los ginkgos retienen varios caracteres de las espermatófitas ancestrales, como el polen que emite un tubo haustorial (no polínico), y el esperma flagelado capaz de nadar.
Son árboles o arbustos con madera bien desarrollada y usualmente hojas de tipo aguja.
Normalmente las hojas son solitarias, naciendo a lo largo del tallo, pero en los pinos (Pinus) están agrupadas en ramitas pequeñas.
Los datos moleculares muestran una división basal entre las pináceas y un clado que albergaría a todas las demás coníferas.
Las pináceas poseen varias características singulares, como óvulos invertidos (con la micropila mirando hacia el eje del cono) y las semillas aladas, alas que se originan en la escama ovulífera durante el desarrollo de la semilla.
Las cupresáceas están marcadas por muchas características singulares, como la fusión de la escama ovulífera con la báctea tectriz.
Este grupo contiene solo unas 80 especies vivientes, que pertenecen a tres linajes bastante diferenciados (Doyle 1996, Friedman 1996, Price 1996).
Finalmente, Welwitschia con una sola especie, Welwitschia mirabilis encontrada en el sudoeste de África, produce solo dos o raramente cuatro hojas funcionales a lo largo de toda su vida, hojas que crecen indefinidamente por meristemas presentes en la base, y se necrosan gradualmente en las puntas.
Si bien estos tres clados se ven muy diferentes uno del otro, comparten muchas características inusuales, como las hojas opuestas, múltiples yemas por axila, vasos xilemáticos con aberturas circulares entre células adjuntas, polen compuesto, semillas en estróbilo, y un polen elipsoide ancestral con unas estrías características que corren de punta a punta.
En marcado contraste con las angiospermas en cambio, las gnetofitas nunca se volvieron un componente significativo de la flora en paleolatitudes altas y medias, y han sufrido una disminución dramática de su representatividad durante el Cretácico tardío (Crane et al.

Referencias:
n : generación haploide,
2n : generación diploide,
m! : mitosis,
M! : meiosis,
F! : fecundación