Un plásmido inductor de tumores (Ti) es un plásmido que se encuentra en especies patógenas de Agrobacterium , incluidas A. tumefaciens , A. rhizogenes , A. rubi y A. vitis .
Evolutivamente, el plásmido Ti es parte de una familia de plásmidos que portan muchas especies de Alphaproteobacteria . Los miembros de esta familia de plásmidos se definen por la presencia de una región de ADN conservada conocida como casete del gen repABC , que media la replicación del plásmido, la partición del plásmido en células hijas durante la división celular , así como el mantenimiento del plásmido en números bajos de copias en una célula. [1] Los plásmidos Ti en sí mismos se clasifican en diferentes categorías según el tipo de molécula, o bien , permiten que las bacterias se descompongan como fuente de energía. [2]
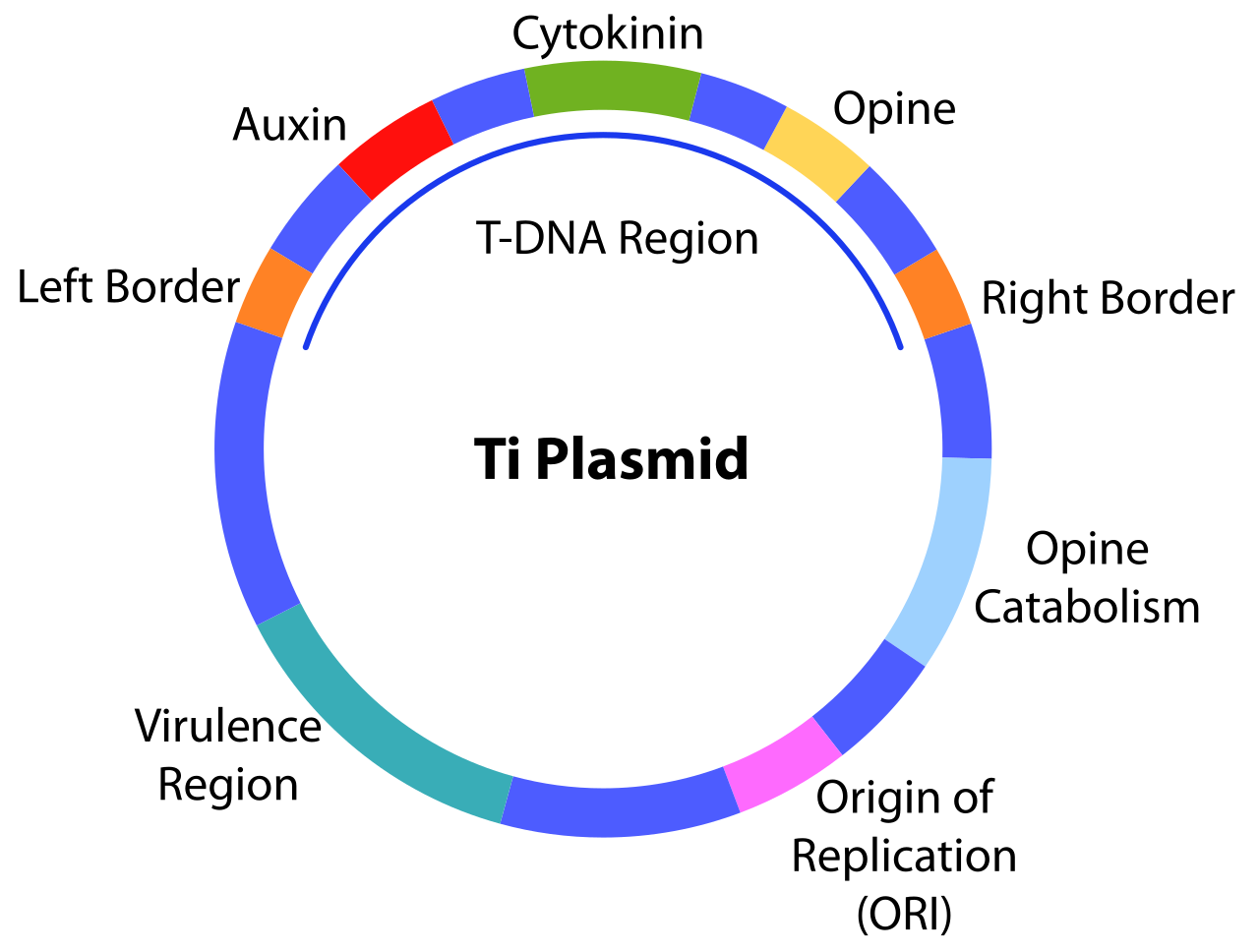
La presencia de este plásmido Ti es esencial para que las bacterias causen la enfermedad de la agalla de la corona en las plantas. [1] Esto se facilita a través de ciertas regiones cruciales en el plásmido Ti, incluida la región vir , que codifica genes de virulencia, y la región de ADN de transferencia (T-ADN), que es una sección del plásmido Ti que se transfiere mediante conjugación a las células de la planta huésped después de que la bacteria detecta un sitio de lesión. Estas regiones tienen características que permiten la entrega de T-ADN a las células de la planta huésped y pueden modificar la célula de la planta huésped para causar la síntesis de moléculas como hormonas vegetales (por ejemplo, auxinas , citoquininas ) y opinas y la formación de tumores de agalla de la corona. [1]
Debido a que la región T-ADN del plásmido Ti puede transferirse de bacterias a células vegetales, representó una vía interesante para la transferencia de ADN entre reinos y estimuló grandes cantidades de investigación sobre el plásmido Ti y sus posibles usos en bioingeniería.
El plásmido Ti es un miembro de la familia de plásmidos RepABC que se encuentra en Alphaproteobacteria. [3] Estos plásmidos suelen ser de tamaño relativamente grande, desde 100 kbp hasta 2 Mbp. También se los suele denominar replicones , ya que su replicación comienza en un solo sitio. Los miembros de esta familia tienen un casete genético repABC característico. [4] Otro miembro notable de esta familia es el plásmido inductor de raíces (Ri) transportado por A. rhizogenes , que causa otra enfermedad de las plantas conocida como enfermedad de la raíz pilosa. [1]
Una característica clave de los plásmidos Ti es su capacidad de impulsar la producción de opinas, que son derivados de varios aminoácidos o fosfatos de azúcar , en las células vegetales hospedantes. Estas opinas pueden luego ser utilizadas como nutriente para las bacterias infectantes, que catabolizan las respectivas opinas utilizando genes codificados en el plásmido Ti.
En consecuencia, los plásmidos Ti se han clasificado en función del tipo de opina que catabolizan, a saber: tipos nopalina , octopina o manitilo, que son derivados de aminoácidos, o tipo agrocinopina, que son derivados de fosfato de azúcar. [1]
La identificación de A. tumefaciens como la causa de los tumores biliares en las plantas allanó el camino para comprender la base molecular de la enfermedad de las agallas de la corona. [5]
El primer indicio de un efecto genético en las células de las plantas hospedadoras se produjo en 1942-1943, cuando se descubrió que las células vegetales de tumores secundarios carecían de células bacterianas en su interior. Sin embargo, estas células tumorales sí poseían la capacidad de producir opinas metabolizadas por la cepa bacteriana infectante. [6] Fundamentalmente, la producción de las respectivas opinas se produjo independientemente de la especie de planta y, ocasionalmente, solo dentro de los tejidos de las agallas de la corona, lo que indica que las bacterias habían transferido algún material genético a las células vegetales hospedadoras para permitir la síntesis de opinas. [5]
Sin embargo, la forma y el grado en que se produjo la transferencia de ADN sigue siendo una pregunta abierta. La adición de ADN de A. tumefaciens por sí sola no provocó tumores en las plantas [7] , mientras que se encontró que muy poco ADN de A. tumefaciens se integraba en el genoma de la célula vegetal huésped [8] . La adición de desoxirribonucleasas (DNasas) para degradar el ADN tampoco logró evitar la formación y el crecimiento de los tumores de la planta [9] . Estos sugirieron que poco, si es que algo, del ADN de A. tumefaciens se transfiere a la célula vegetal huésped para causar enfermedades y, si el ADN se transfiere de hecho de la bacteria a la planta, debe ocurrir de manera protegida.
Posteriormente, se descubrió que las cepas bacterianas oncogénicas podían convertir bacterias no patógenas en patógenas mediante el proceso de conjugación, donde los genes responsables de la virulencia se transferían a las células no patógenas. [10] El papel de un plásmido en esta capacidad patógena se respaldó aún más cuando se encontraron plásmidos grandes solo en bacterias patógenas pero no en bacterias avirulentas. [11] Finalmente, se estableció la detección de partes de plásmidos bacterianos en células de plantas hospedadoras, lo que confirmó que este era el material genético responsable del efecto genético de la infección. [12]
Con la identificación del plásmido Ti, se llevaron a cabo muchos estudios para determinar las características del plásmido Ti y cómo se transfiere el material genético de Agrobacterium a la planta huésped. Algunos hitos notables en los estudios de plásmidos Ti incluyen el mapeo de un plásmido Ti en 1978 y el estudio de la similitud de secuencias entre diferentes plásmidos Ti en 1981. [13] [14]
Entre 1980 y 2000, también se llevó a cabo la caracterización de la región T-ADN y la región "vir". Los estudios sobre la región T-ADN determinaron su proceso de transferencia e identificaron genes que permiten la síntesis de hormonas vegetales y opinas. [15] Por otra parte, los primeros trabajos apuntaron a determinar las funciones de los genes codificados en la región "vir", que se clasificaron en general en aquellos que permitían interacciones entre bacterias y huéspedes y aquellos que posibilitaban la administración de T-ADN. [2]

La replicación, partición y mantenimiento del plásmido Ti depende del casete del gen repABC , que se compone principalmente de tres genes: repA , repB y repC . repA y repB codifican cada uno proteínas implicadas en la partición del plásmido, mientras que repC codifica un iniciador de la replicación. [1] Estos genes se expresan a partir de 4 promotores diferentes ubicados aguas arriba de repA . repE codifica un pequeño ARN antisentido y se encuentra entre repB y repC . [4] Además, hay un sitio de partición ( parS ) y un origen de replicación ( oriV ) presentes dentro del casete repABC . [1]
La replicación del plásmido Ti es impulsada por la proteína iniciadora RepC ( P05684 ), que posee dos dominios proteicos : un dominio N-terminal (NTD) que se une al ADN y un dominio C-terminal (CTD). Los análisis mutacionales han demostrado que sin una proteína RepC funcional, el plásmido Ti no puede replicarse. [4] Mientras tanto, la secuencia oriV tiene alrededor de 150 nucleótidos de longitud y se encuentra dentro del gen repC . [3] Los experimentos de laboratorio han demostrado que la proteína RepC se une a esta región, lo que sugiere su papel como origen de la replicación. [16] Por lo tanto, aunque el proceso completo detrás de la replicación del plásmido Ti no se ha descrito por completo, el paso inicial de la replicación probablemente dependería de la expresión de RepC y su unión a oriV . Cabe destacar que la proteína RepC solo actúa en cis , donde solo impulsa la replicación del plásmido en el que está codificada y no de ningún otro plásmido también presente en la célula bacteriana. [16]
El sistema de partición del plásmido Ti es similar al sistema ParA/ParB utilizado en otros plásmidos y cromosomas bacterianos y se cree que actúa de la misma manera. [17] Las mutaciones en cualquiera de las proteínas RepA o RepB han dado como resultado una disminución en la estabilidad del plásmido, lo que indica su papel e importancia en la partición del plásmido. [4] La capacidad de RepA para formar filamentos le permite crear un puente físico a lo largo del cual el ADN puede ser tirado hacia polos opuestos de una célula en división. Mientras tanto, la proteína RepB puede unirse específicamente a la secuencia parS , formando un complejo con el ADN que puede ser reconocido por RepA. [1] [4] Este sistema es particularmente importante para la partición adecuada del plásmido Ti, ya que el plásmido solo está presente en unos pocos números de copias en la célula bacteriana.
El plásmido Ti se mantiene en un bajo número de copias dentro de una célula bacteriana. Esto se logra en parte influyendo en la expresión del iniciador de replicación RepC. [1] Cuando se une a ADP , RepA se activa para trabajar con RepB, actuando como un regulador negativo del casete repABC . [3] Por lo tanto, los niveles de RepC se mantienen bajos dentro de una célula, lo que evita que se produzcan demasiadas rondas de replicación durante cada ciclo de división celular. Además, hay un pequeño ARN conocido como RepE codificado entre repB y repC que reduce la expresión de repC . [18] RepE es complementario a RepC y se unirá al ARNm de repC para formar una molécula de doble cadena. Esto puede bloquear la producción traduccional de la proteína RepC. [18]
Por otra parte, la expresión del casete repABC y, por lo tanto, el número de copias del plásmido Ti también se ve influenciada por un sistema de detección de quórum en Agrobacterium . [4] Los sistemas de detección de quórum responden a las densidades de población bacteriana detectando una molécula, conocida como autoinductor, que es producida por las células bacterianas en niveles bajos y que se acumularía hasta un nivel umbral cuando hay una alta densidad de bacterias presentes. [18] En este caso, el autoinductor es la molécula N-3-oxooctanoil-L-homoserina lactona (3-OC 8 -AHL), que es detectada por un regulador conocido como TraR. [4] Cuando se activa, TraR se unirá a regiones conocidas como cajas tra en las regiones promotoras del casete del gen repABC para impulsar la expresión. Por lo tanto, un alto nivel de densidad de población aumenta el número de plásmidos presentes dentro de cada célula bacteriana, lo que probablemente respalde la patogénesis en la planta huésped. [4]

La expresión de la región vir suele estar reprimida en condiciones normales y solo se activa cuando la bacteria detecta señales derivadas de la planta provenientes de los sitios de la herida. Esta activación es necesaria para la producción de proteínas Vir y la transferencia de ADN y proteínas a las células de la planta huésped. [1]
VirA y VirG forman un sistema regulador de dos componentes dentro de Agrobacterium . [19] Este es un tipo de sistema de detección y señalización que se encuentra comúnmente en bacterias; en este caso, actúan para detectar señales derivadas de plantas para impulsar la expresión de la región vir . Durante la detección, VirA, una quinasa sensora de histidina, se fosforilará antes de pasar este grupo fosfato al regulador de respuesta VirG. [20] El regulador de respuesta activado VirG puede luego unirse a una región de ADN conocida como caja vir , ubicada aguas arriba de cada promotor vir , para activar la expresión de la región vir . [1] [19] Una posible función descendente de la detección mediada por VirA y VirG es el movimiento direccional, o quimiotaxis , de las bacterias hacia señales derivadas de plantas; esto permite que Agrobacterium se mueva hacia el sitio de la herida en las plantas. [21] Además, con la inducción de la región vir , la transferencia de T-ADN puede ser mediada por las proteínas Vir. [22]
El operón virB es el operón más grande de la región vir y codifica 11 proteínas VirB involucradas en el proceso de transferencia de ADN-T y proteínas bacterianas a las células de la planta huésped (ver el aparato de transferencia a continuación). [23] [24]
El operón virC codifica dos proteínas: VirC1 y VirC2. Estas proteínas influyen en la patogénesis de Agrobacterium hacia diferentes huéspedes vegetales, y las mutaciones pueden reducir pero no eliminar la virulencia de las bacterias. [25] Tanto los operones virC como virD pueden ser reprimidos por una proteína codificada cromosómicamente conocida como Ros. [26] [27] Ros se une a una región de ADN que se superpone con el sitio de unión del regulador VirG, y por lo tanto compite con VirG para controlar sus niveles de expresión. [26] [27] Funcionalmente, VirC1 y VirC2 promueven el ensamblaje de un complejo relaxosoma durante la transferencia conjugativa de T-ADN de la bacteria a la célula vegetal huésped. [28] Este es un proceso dependiente de la energía mediado a través de su actividad NTPasa, y ocurre cuando se unen a una región de ADN conocida como overdrive . [28] Como resultado, actúan para aumentar la cantidad de hebras de T-ADN producidas. Después de la producción de la cadena de ADN que se va a transferir (cadena de transferencia, cadena T), las proteínas VirC también pueden ayudar a dirigir la cadena de transferencia al aparato de transferencia. [28]
El operón virD codifica para 4 proteínas: VirD1-D4. [29] VirD1 y VirD2 están involucrados en el procesamiento de T-ADN durante la conjugación para producir la cadena T; esta es la molécula de ADN monocatenario que se transporta a la célula vegetal huésped (ver aparato de transferencia a continuación). [30] Durante el procesamiento, VirD1 actuará como una topoisomerasa para desenrollar las cadenas de ADN. [30] VirD2, una relaxasa , luego cortará una de las cadenas de ADN y permanecerá unida al ADN mientras se transfiere a la célula receptora. [31] [32] Dentro de la célula receptora, VirD2 también trabajará junto con VirE2 para dirigir el ADN transferido al núcleo de la célula receptora. Hay sugerencias de que VirD2 puede ser fosforilado y desfosforilado por diferentes proteínas, lo que afecta su capacidad para entregar ADN. [33] Por el contrario, se sabe poco sobre VirD3, y los análisis mutacionales no han proporcionado ningún respaldo a su papel en la virulencia de Agrobacterium . [34] Finalmente, VirD4 es una parte crucial del proceso de conjugación, ya que actúa como un factor de acoplamiento que reconoce y transfiere la cadena T al canal de transporte. [35]
El operón virE codifica para 2 proteínas: VirE1 y VirE2. [36] VirE2 es una proteína efectora translocada junto con la hebra T a las células de la planta huésped. Allí, se une a la hebra T para dirigir su entrega al núcleo de la célula vegetal huésped. [37] [38] Parte de esta actividad implica la presencia de secuencias de localización nuclear dentro de la proteína, que marca la proteína y el ADN asociado para la entrada al núcleo. También protege la hebra T del ataque de la nucleasa . [39] Existe cierta especulación con respecto al papel de VirE2 como un canal de proteína, permitiendo que el ADN se mueva a través de la membrana citoplasmática de la planta . [40] Por otro lado, VirE1 puede estar involucrado en la promoción de la transferencia de la proteína VirE2 a la célula vegetal huésped. [41] Se une al dominio de unión a ssDNA de VirE2, evitando así que la proteína VirE2 se una prematuramente a la hebra T dentro de la célula bacteriana. [42]
virF es un factor de especificidad del hospedador que se encuentra en algunos, pero no en todos los tipos de plásmidos Ti; por ejemplo, los plásmidos Ti de tipo octopina poseen virF pero los de tipo nopalina no. [43] [44] La capacidad de A. tumefaciens para inducir tumores de agallas de corona en ciertas especies de plantas pero no en otras se ha atribuido a la presencia o ausencia de este gen virF . [43] [44]
El operón virH codifica dos proteínas: VirH1 y VirH2. [45] Un estudio bioinformático de las secuencias de aminoácidos de la proteína VirH mostró similitudes entre ellas y una superfamilia de proteínas conocidas como enzimas del citocromo P450 . [46] Luego se descubrió que VirH2 metaboliza ciertos compuestos fenólicos detectados por VirA. [45]
El ADN-T de Agrobacterium tiene una longitud de aproximadamente 15-20 kbp y se integrará en el genoma de la planta huésped tras su transferencia a través de un proceso conocido como recombinación . Este proceso utiliza huecos preexistentes en el genoma de la célula vegetal huésped para permitir que el ADN-T se empareje con secuencias cortas en el genoma, lo que prepara el proceso de ligación del ADN , donde el ADN-T se une permanentemente al genoma de la planta. [37] La región del ADN-T está flanqueada en ambos extremos por secuencias de 24 pb.
Dentro del genoma de la célula vegetal huésped, el ADN-T de Agrobacterium se expresa para producir dos grupos principales de proteínas. [1] Un grupo es responsable de la producción de hormonas de crecimiento de las plantas. A medida que se producen estas hormonas, habrá un aumento en la tasa de división celular y, por lo tanto, la formación de tumores de agallas de la corona. [47] El segundo grupo de proteínas es responsable de impulsar la síntesis de opinas en las células vegetales huésped. Las opinas específicas producidas dependen del tipo de plásmido Ti, pero no de la planta huésped. Estas opinas no pueden ser utilizadas por la planta huésped y, en su lugar, se exportarán fuera de la célula vegetal, donde pueden ser absorbidas por las células de Agrobacterium . Las bacterias poseen genes en otras regiones del plásmido Ti que permiten el catabolismo de las opinas. [1]
Los aparatos de transferencia codificados dentro del plásmido Ti tienen que lograr dos objetivos: permitir la transferencia conjugativa del plásmido Ti entre bacterias y permitir la entrega del T-ADN y ciertas proteínas efectoras a las células de la planta huésped. Esto se logra mediante el sistema Tra/Trb y el sistema VirB/VirD4 respectivamente, que son miembros del sistema de secreción tipo IV (T4SS). [47]
Para que el plásmido Ti y el ADN-T se transfieran mediante conjugación, primero deben ser procesados por diferentes proteínas, como la enzima relaxasa (TraA/VirD2) y las proteínas de transferencia y replicación del ADN (Dtr). Juntas, estas proteínas reconocerán y se unirán a una región conocida como el origen de transferencia ( oriT ) en el plásmido Ti para formar el complejo relaxosoma. Para el ADN-T, se creará una muesca en la secuencia límite del ADN-T, y la hebra T mellada se transportará a la membrana celular, donde está presente el resto de la maquinaria de transferencia. [31]
Dentro del sistema VirB/VirD4, la relaxasa VirD2 es asistida por los factores accesorios VirD1, VirC1 y VirC2 mientras procesa el sustrato de ADN. [48] Además, la relaxasa VirD2 y las proteínas VirC contribuirán a la entrega de la cadena de ADN al receptor VirD4 en la membrana celular. [28] Este receptor es un componente esencial de los T4SS y se cree que energiza y media la transferencia del ADN al canal de translocación entre dos células. [49] La siguiente tabla resume las proteínas codificadas en el operón virB que compone el canal de translocación del sistema VirB/VirD4. [1]
La capacidad de Agrobacterium para introducir ADN en las células vegetales abrió nuevas puertas para la ingeniería genómica vegetal , permitiendo la producción de plantas modificadas genéticamente (plantas transgénicas). [57] Las proteínas implicadas en la mediación de la transferencia de T-ADN reconocerán primero las secuencias limítrofes de la región de T-ADN. Por lo tanto, es posible que los científicos utilicen secuencias limítrofes de T-ADN para flanquear cualquier secuencia de interés deseada; dicho producto puede insertarse entonces en un plásmido e introducirse en las células de Agrobacterium . [58] Allí, las secuencias limítrofes serán reconocidas por el aparato de transferencia de A. tumefaciens y se introducirán de forma estándar en la célula vegetal de destino. [1] Además, al dejar atrás solo las secuencias limítrofes del T-ADN, el producto resultante editará el genoma de la planta sin causar ningún tumor en las plantas. [59] Este método se ha utilizado para modificar varias plantas de cultivo, entre ellas el arroz, [60] la cebada [61] y el trigo. [62] Desde entonces, trabajos posteriores han ampliado los objetivos de A. tumefaciens para incluir hongos y líneas celulares humanas. [63] [64]