
El impacto humano en el ciclo del nitrógeno es diverso. Los aportes de nitrógeno (N) agrícola e industrial al medio ambiente actualmente superan los aportes provenientes de la fijación natural de N. [1] Como consecuencia de los aportes antropogénicos, el ciclo global del nitrógeno (Fig. 1) se ha alterado significativamente durante el último siglo. Las fracciones molares atmosféricas globales de óxido nitroso (N 2 O) han aumentado de un valor preindustrial de ~270 nmol/mol a ~319 nmol/mol en 2005. [2] Las actividades humanas representan más de un tercio de las emisiones de N 2 O , la mayor parte de los cuales se deben al sector agrícola. [2] Este artículo tiene como objetivo ofrecer una breve revisión de la historia de los aportes antropogénicos de N y los impactos reportados de los aportes de nitrógeno en ecosistemas terrestres y acuáticos seleccionados .
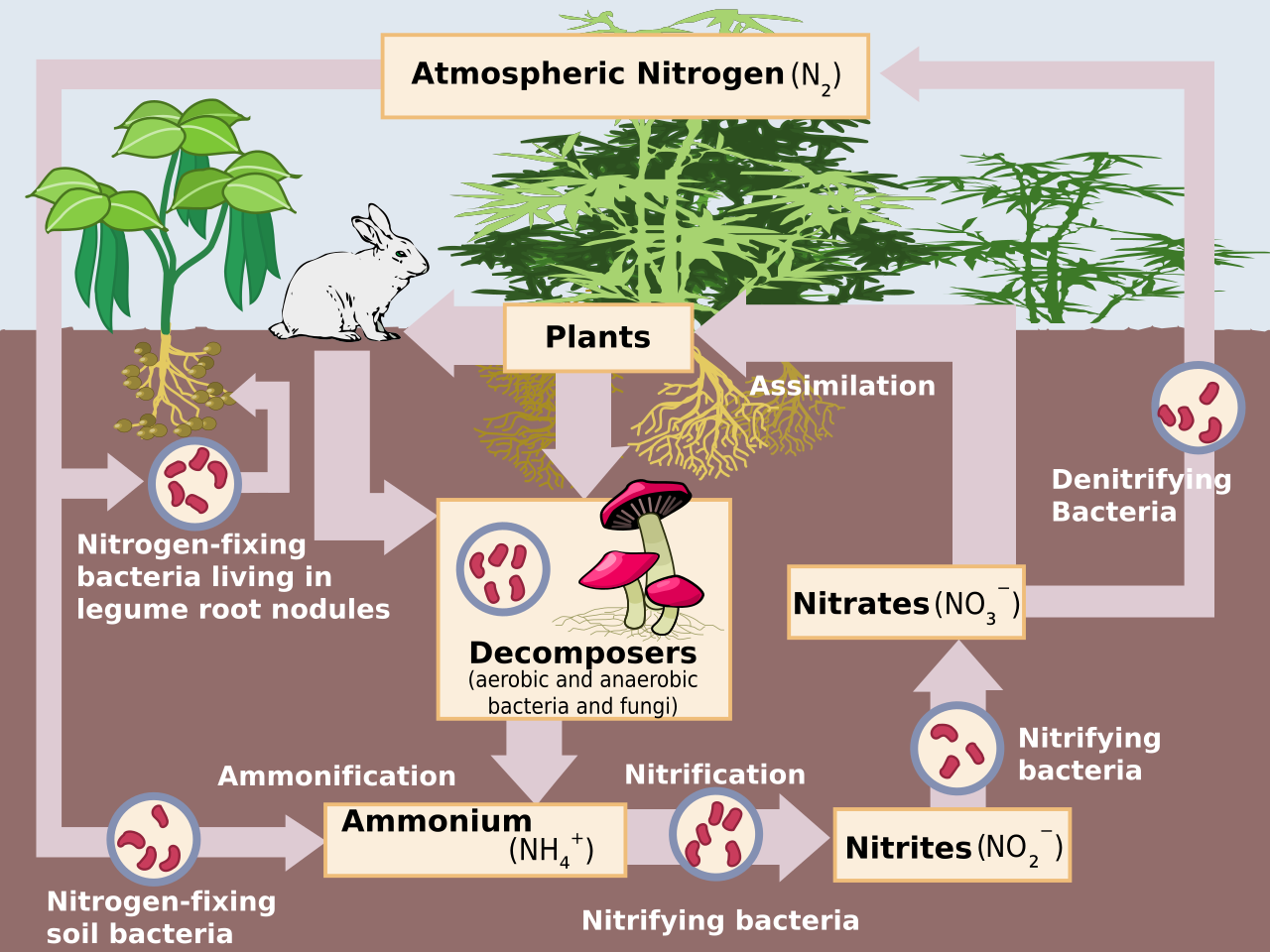
Aproximadamente el 78% de la atmósfera de la Tierra es gas N (N 2 ), que es un compuesto inerte y biológicamente no disponible para la mayoría de los organismos. Para poder ser utilizado en la mayoría de los procesos biológicos, el N 2 debe convertirse en nitrógeno reactivo (Nr), que incluye formas inorgánicas reducidas (NH 3 y NH 4 + ), formas inorgánicas oxidadas (NO, NO 2 , HNO 3 , N 2 O, y NO 3 − ), y compuestos orgánicos ( urea , aminas y proteínas ). [1] El N 2 tiene un triple enlace fuerte, por lo que se requiere una cantidad significativa de energía (226 kcal mol −1 ) para convertir N 2 en Nr. [1] Antes de los procesos industriales, las únicas fuentes de dicha energía eran la radiación solar y las descargas eléctricas. [1] Utilizando una gran cantidad de energía metabólica y la enzima nitrogenasa , algunas bacterias y cianobacterias convierten el N 2 atmosférico en NH 3 , un proceso conocido como fijación biológica de nitrógeno (BNF). [4] El análogo antropogénico del BNF es el proceso de Haber-Bosch , en el que el H 2 reacciona con el N 2 atmosférico a altas temperaturas y presiones para producir NH 3 . [5] Por último, el N 2 se convierte en NO mediante la energía procedente de los rayos , que es insignificante en los ecosistemas templados actuales, o mediante la quema de combustibles fósiles . [1]
Hasta 1850, el BNF natural, el BNF inducido por el cultivo (por ejemplo, la plantación de leguminosas ) y la materia orgánica incorporada eran las únicas fuentes de N para la producción agrícola. [5] Hacia principios de siglo, el Nr de los depósitos de guano y nitrato de sodio se recolectaba y exportaba desde las áridas islas del Pacífico y los desiertos de América del Sur. [5] A finales de la década de 1920, los primeros procesos industriales, aunque ineficaces, se utilizaban habitualmente para producir NH 3 . [1] Gracias a los esfuerzos de Fritz Haber y Carl Bosch , el proceso Haber-Bosch se convirtió en la mayor fuente de fertilizantes nitrogenados después de la década de 1950 y reemplazó al BNF como fuente dominante de producción de NH 3 . [5] De 1890 a 1990, el Nr creado antropogénicamente aumentó casi nueve veces. [1] Durante este tiempo, la población humana se triplicó, en parte debido al aumento de la producción de alimentos.
Desde la revolución industrial , una fuente adicional de aporte antropogénico de N ha sido la quema de combustibles fósiles , que se utiliza para liberar energía (por ejemplo, para impulsar automóviles). A medida que se queman combustibles fósiles , las altas temperaturas y presiones proporcionan energía para producir NO a partir de la oxidación del N 2 . [1] Además, cuando se extrae y quema combustible fósil , el N fósil puede volverse reactivo (es decir, emisiones de NOx ). [1] Durante la década de 1970, los científicos comenzaron a reconocer que los aportes de N se estaban acumulando en el medio ambiente y afectando a los ecosistemas. [1]
Entre 1600 y 1990, la creación global de nitrógeno reactivo (Nr) había aumentado casi un 50%. [6] Durante este período, las emisiones atmosféricas de especies de Nr supuestamente aumentaron un 250% y la deposición en los ecosistemas marinos y terrestres aumentó más del 200%. [6] Además, se informó que se cuadruplicó el flujo de N inorgánico disuelto en los ríos hacia las costas. [6] El nitrógeno es un nutriente limitante crítico en muchos sistemas, incluidos bosques, humedales y ecosistemas costeros y marinos; por lo tanto, este cambio en las emisiones y distribución de Nr ha tenido consecuencias sustanciales para los ecosistemas acuáticos y terrestres. [7] [8]
Las entradas de N atmosférico incluyen principalmente óxidos de N (NO x ), amoníaco (NH 3 ) y óxido nitroso (N 2 O) de ecosistemas acuáticos y terrestres, [4] y NO x de la combustión de combustibles fósiles y biomasa. [1]
En los agroecosistemas , la aplicación de fertilizantes ha aumentado la nitrificación microbiana (proceso aeróbico en el que los microorganismos oxidan el amonio [NH 4 + ] a nitrato [NO 3 − ]) y la desnitrificación (proceso anaeróbico en el que los microorganismos reducen el NO 3 − al gas nitrógeno atmosférico [N 2 ] ). Ambos procesos liberan naturalmente óxido nítrico (NO) y óxido nitroso (N 2 O) a la atmósfera. [4] De particular preocupación es el N 2 O, que tiene una vida atmosférica promedio de 114 a 120 años, [10] y es 300 veces más efectivo que el CO 2 como gas de efecto invernadero . [4] El NOx producido por procesos industriales, automóviles y fertilización agrícola y el NH3 emitido por los suelos (es decir, como subproducto adicional de la nitrificación) [4] y las operaciones ganaderas se transportan a ecosistemas a favor del viento, lo que influye en el ciclo del N y las pérdidas de nutrientes. Se han citado seis efectos principales de las emisiones de NOx y NH3 : [1] 1) disminución de la visibilidad atmosférica debido a los aerosoles de amonio ( partículas finas [PM]); 2) concentraciones elevadas de ozono ; 3) el ozono y las partículas afectan la salud humana (por ejemplo, enfermedades respiratorias , cáncer ); 4) aumentos en el forzamiento radiativo y el cambio climático global ; 5) disminución de la productividad agrícola debido a la deposición de ozono ; y 6) acidificación de los ecosistemas [11] y eutrofización .
Los ecosistemas terrestres y acuáticos reciben aportes de Nr de la atmósfera a través de deposición húmeda y seca. [1] Las especies de Nr atmosférico pueden depositarse en los ecosistemas en forma de precipitación (p. ej., NO 3 − , NH 4 + , compuestos orgánicos de N), como gases (p. ej., NH 3 y ácido nítrico gaseoso [HNO 3 ]) o como aerosoles ( ej., nitrato de amonio [NH 4 NO 3 ]). [1] Los ecosistemas acuáticos reciben nitrógeno adicional de la escorrentía superficial y los aportes fluviales . [8]
El aumento de la deposición de N puede acidificar suelos, arroyos y lagos y alterar la productividad de bosques y pastizales. En los ecosistemas de pastizales, los aportes de N han producido aumentos iniciales en la productividad seguidos de disminuciones a medida que se exceden los umbrales críticos. [1] También se han demostrado los efectos del nitrógeno sobre la biodiversidad , el ciclo del carbono y los cambios en la composición de las especies . En áreas altamente desarrolladas de sistemas costeros oceánicos y estuarinos cercanos a la costa, los ríos suministran aportes de N directos (p. ej., escorrentía superficial ) e indirectos (p. ej., contaminación de las aguas subterráneas) provenientes de los agroecosistemas. [8] El aumento de los aportes de N puede provocar la acidificación del agua dulce y la eutrofización de las aguas marinas.
Gran parte del crecimiento terrestre en los sistemas templados está limitado por el N; por lo tanto, los aportes de N (es decir, a través de la deposición y la fertilización) pueden aumentar la disponibilidad de N, lo que aumenta temporalmente la absorción de N, el crecimiento vegetal y microbiano, y la acumulación de N en la biomasa vegetal y la materia orgánica del suelo . [12] La incorporación de mayores cantidades de N en la materia orgánica disminuye las proporciones C:N, aumentando la liberación de N mineral (NH 4 + ) durante la descomposición de la materia orgánica por microbios heterótrofos (es decir, amonificación ). [13] A medida que aumenta la amonificación, también aumenta la nitrificación del N mineralizado. Debido a que la nitrificación y desnitrificación microbianas tienen "fugas", se espera que la deposición de N aumente las emisiones de gases traza. [14] Además, con el aumento de la acumulación de NH 4 + en el suelo, los procesos de nitrificación liberan iones de hidrógeno, que acidifican el suelo. El NO 3 − , el producto de la nitrificación, es muy móvil y puede lixiviarse del suelo, junto con minerales alcalinos cargados positivamente, como el calcio y el magnesio. [4] En suelos ácidos, los iones de aluminio movilizados pueden alcanzar concentraciones tóxicas, afectando negativamente tanto a los ecosistemas terrestres como a los acuáticos adyacentes.
Las fuentes antropogénicas de N generalmente llegan a los bosques de tierras altas mediante deposición . [15] Una posible preocupación por el aumento de la deposición de N debido a las actividades humanas es la alteración del ciclo de nutrientes en los ecosistemas forestales. Numerosos estudios han demostrado los impactos tanto positivos como negativos de la deposición de N atmosférico en la productividad forestal y el almacenamiento de carbono. El N añadido suele ser rápidamente inmovilizado por los microbios [ 16] y el efecto del N restante disponible depende de la capacidad de la comunidad vegetal para absorber N. [17] En sistemas con alta absorción, el N se asimila a la biomasa vegetal, lo que conduce a una mayor productividad primaria neta (PPN) y posiblemente a un mayor secuestro de carbono a través de una mayor capacidad fotosintética. Sin embargo, las respuestas de los ecosistemas a las adiciones de N dependen de muchos factores específicos del sitio, incluido el clima, el historial de uso de la tierra y la cantidad de adiciones de N. Por ejemplo, en el noreste de Estados Unidos, los rodales de frondosas que reciben aportes crónicos de N han demostrado una mayor capacidad para retener N y aumentar la productividad primaria neta anual (ANPP) que los rodales de coníferas. [18] Una vez que la entrada de N excede la demanda del sistema, el N puede perderse a través de lixiviación y flujos de gas. Cuando el N disponible excede la capacidad de absorción del ecosistema (es decir, la vegetación, el suelo y los microbios , etc.), se produce la saturación de N y el exceso de N se pierde en las aguas superficiales, las aguas subterráneas y la atmósfera. [12] [17] [18] La saturación de N puede provocar desequilibrios de nutrientes (p. ej., pérdida de calcio debido a la lixiviación de nitratos) y una posible disminución de los bosques. [13]
Un estudio de 15 años de duración sobre las adiciones crónicas de N en el programa de Investigación Ecológica a Largo Plazo Forestal ( LTER ) de Harvard ha dilucidado muchos impactos del aumento de la deposición de nitrógeno en el ciclo de nutrientes en los bosques templados. Encontró que las adiciones crónicas de N resultaban en mayores pérdidas por lixiviación, mayor mortalidad de los pinos y cese de la acumulación de biomasa. [18] Otro estudio informó que las adiciones crónicas de N dieron como resultado la acumulación de N no fotosintético y posteriormente redujeron la capacidad fotosintética, lo que supuestamente condujo a un estrés severo por carbono y mortalidad. [17] Estos hallazgos niegan hipótesis anteriores de que un aumento en los aportes de N aumentaría la PNP y el secuestro de carbono .
Muchas comunidades de plantas han evolucionado en condiciones de escasez de nutrientes; por lo tanto, un aumento en los aportes de N puede alterar las interacciones bióticas y abióticas, lo que lleva a cambios en la composición de la comunidad. Varios estudios de adición de nutrientes han demostrado que el aumento de los aportes de N conduce a la dominancia de especies de plantas de rápido crecimiento, con la consiguiente disminución de la riqueza de especies. [19] [20] [21] Las especies de crecimiento rápido tienen una mayor afinidad por la absorción de nitrógeno y desplazarán a las especies de plantas de crecimiento más lento al bloquear el acceso a la luz solar con su mayor biomasa sobre el suelo. [22] Otros estudios han encontrado que las respuestas secundarias del sistema al enriquecimiento de N, incluida la acidificación del suelo y los cambios en las comunidades de micorrizas , han permitido que las especies tolerantes al estrés superen a las especies sensibles. [11] [23] Los árboles que tienen asociaciones de micorrizas arbusculares tienen más probabilidades de beneficiarse de un aumento en el nitrógeno del suelo, ya que estos hongos no pueden descomponer el nitrógeno orgánico del suelo. [24] Otros dos estudios encontraron evidencia de que el aumento de la disponibilidad de N ha resultado en disminuciones en los brezales con especies diversas . Los brezales se caracterizan por suelos pobres en N, que excluyen los pastos exigentes en N; sin embargo, con el aumento de la deposición de N y la acidificación del suelo , los pastizales invasores reemplazan a los brezales de las tierras bajas. [25] [26]
En un estudio experimental más reciente sobre fertilización con N y perturbación (es decir, labranza) en la sucesión de campos antiguos, se encontró que la riqueza de especies disminuía al aumentar el N, independientemente del nivel de perturbación. Los experimentos de competencia mostraron que los dominantes competitivos excluían a las especies competitivamente inferiores entre eventos de perturbación. Con mayores aportes de N, la competencia pasó del subsuelo a la superficie (es decir, a la competencia por la luz) y las tasas de colonización de parches disminuyeron significativamente. Estos cambios internos pueden afectar dramáticamente a la comunidad al cambiar el equilibrio de las compensaciones entre competencia y colonización entre especies. [21] En los sistemas basados en parches, la coexistencia regional puede ocurrir a través de compensaciones en capacidades competitivas y colonizadoras dadas tasas de perturbación suficientemente altas. [27] Es decir, con una clasificación inversa de capacidades competitivas y colonizadoras, las plantas pueden coexistir en el espacio y el tiempo a medida que las perturbaciones eliminan a los competidores superiores de los parches, lo que permite el establecimiento de colonizadores superiores. Sin embargo, como lo demostraron Wilson y Tilman, el aumento de los aportes de nutrientes puede anular las compensaciones, lo que resulta en la exclusión competitiva de estos colonizadores superiores/competidores pobres. [21]
Los ecosistemas acuáticos también exhiben respuestas variadas al enriquecimiento de nitrógeno. NO 3 : la carga de ecosistemas terrestres saturados de N puede provocar la acidificación de los sistemas de agua dulce aguas abajo y la eutrofización de los sistemas marinos aguas abajo. La acidificación del agua dulce puede causar toxicidad por aluminio y mortalidad en especies de peces sensibles al pH. Debido a que los sistemas marinos generalmente tienen limitaciones de nitrógeno, los aportes excesivos de N pueden provocar la degradación de la calidad del agua debido a la proliferación de algas tóxicas, la deficiencia de oxígeno, la pérdida de hábitat, la disminución de la biodiversidad y las pérdidas de pesquerías. [8]
La deposición de N atmosférico en paisajes terrestres puede transformarse a través de procesos microbianos del suelo en nitrógeno biológicamente disponible, lo que puede resultar en la acidificación de las aguas superficiales y la pérdida de biodiversidad . Los aportes de NO 3 − y NH 4 + de los sistemas terrestres y de la atmósfera pueden acidificar los sistemas de agua dulce cuando hay poca capacidad amortiguadora debido a la acidificación del suelo . [8] La contaminación por N en Europa, el noreste de Estados Unidos y Asia es una preocupación actual por la acidificación del agua dulce . [28] Los estudios de acidificación de lagos en el Área Experimental del Lago (ELA) en el noroeste de Ontario demostraron claramente los efectos negativos del aumento de la acidez en una especie de pez nativo: el reclutamiento y el crecimiento de la trucha de lago (Salvelinus namaycush) disminuyeron dramáticamente debido a la extirpación de sus principales especies presa. durante la acidificación. [29] El nitrógeno reactivo procedente de la agricultura, la cría de animales, los fertilizantes, los sistemas sépticos y otras fuentes ha elevado las concentraciones de nitrato en las vías fluviales de la mayoría de las naciones industrializadas. Las concentraciones de nitrato en 1.000 lagos noruegos se habían duplicado en menos de una década. Los ríos en el noreste de Estados Unidos y la mayor parte de Europa se han multiplicado entre diez y quince veces durante el último siglo. El nitrógeno reactivo puede contaminar el agua potable a través de su escorrentía hacia arroyos, lagos, ríos y aguas subterráneas. Sólo en los Estados Unidos, hasta el 20% de las fuentes de agua subterránea exceden el límite de concentración de nitrato en agua potable establecido por la Organización Mundial de la Salud. Estas altas concentraciones pueden causar la "enfermedad del bebé azul", donde los iones de nitrato debilitan la capacidad de la sangre para transportar oxígeno. Los estudios también han relacionado las altas concentraciones de nitratos con problemas reproductivos y propensión a algunos cánceres, como el de vejiga y de ovario. [30]
La urbanización, la deforestación y las actividades agrícolas contribuyen en gran medida a la entrada de sedimentos y nutrientes a las aguas costeras a través de los ríos. [8] El aumento de los aportes de nutrientes a los sistemas marinos ha mostrado tanto aumentos a corto plazo en la productividad y el rendimiento pesquero como efectos perjudiciales a largo plazo de la eutrofización . La triplicación de las cargas de NO 3 en el río Mississippi en la última mitad del siglo XX se ha correlacionado con mayores rendimientos pesqueros en las aguas que rodean el delta del Mississippi; [31] sin embargo, estos aportes de nutrientes han producido hipoxia estacional (concentraciones de oxígeno inferiores a 2-3 mg L −1 , " zonas muertas ") en el Golfo de México . [1] [8] En los sistemas estuarinos y costeros, los altos aportes de nutrientes aumentan la producción primaria (por ejemplo, fitoplancton , pastos marinos, macroalgas), lo que aumenta la turbidez con la consiguiente disminución de la penetración de la luz en toda la columna de agua. En consecuencia, el crecimiento de la vegetación sumergida disminuye, lo que reduce la complejidad del hábitat y la producción de oxígeno. El aumento de la producción primaria (es decir, fitoplancton, macroalgas, etc.) conduce a un flujo de carbono hacia las aguas del fondo cuando la materia orgánica en descomposición (es decir, producción primaria senescente) se hunde y es consumida por bacterias aeróbicas que se encuentran más abajo en la columna de agua. Como resultado, el consumo de oxígeno en las aguas del fondo es mayor que la difusión de oxígeno desde las aguas superficiales. Además, ciertas floraciones de algas denominadas floraciones de algas nocivas (FAN) producen toxinas que pueden actuar como compuestos neuromusculares o dañinos para los órganos. Estas floraciones de algas pueden ser perjudiciales para otras formas de vida marina y para los humanos. [32] [33]
Las respuestas del sistema anteriores a las entradas de nitrógeno reactivo (Nr) se estudian casi todas exclusivamente por separado; sin embargo, las investigaciones indican cada vez más que los problemas de carga de nitrógeno están relacionados por múltiples vías que transportan nutrientes a través de los límites del sistema. [1] Esta transferencia secuencial entre ecosistemas se denomina cascada de nitrógeno. [6] (ver ilustración del Programa de las Naciones Unidas para el Medio Ambiente). Durante la cascada, algunos sistemas acumulan Nr, lo que provoca un retraso en la cascada y un aumento de los efectos del Nr en el entorno en el que se acumula. En última instancia, los aportes antropogénicos de Nr se acumulan o se desnitrifican; sin embargo, se ha avanzado poco en la determinación de la importancia relativa de la acumulación y desnitrificación del Nr , lo que se ha debido principalmente a una falta de integración entre las disciplinas científicas. [1] [34]
La mayor parte del Nr se aplica en cascadas de agroecosistemas globales a través de la atmósfera y los ecosistemas acuáticos y terrestres hasta que se convierte en N 2 , principalmente a través de la desnitrificación . [1] Aunque la desnitrificación terrestre produce intermediarios gaseosos (óxido nítrico [NO] y óxido nitroso [N 2 O]), el último paso, la producción microbiana de N 2 , es fundamental porque el N 2 atmosférico es un sumidero de Nr. [34] Muchos estudios han demostrado claramente que las franjas de protección y los humedales gestionados pueden eliminar cantidades significativas de nitrato (NO 3 − ) de los sistemas agrícolas mediante la desnitrificación . [35] Esta gestión puede ayudar a atenuar los efectos en cascada indeseables y eliminar la acumulación ambiental de Nr. [1]
Las actividades humanas dominan los ciclos globales y la mayoría de las regiones del N. [36] Los aportes de N han mostrado consecuencias negativas tanto para el ciclo de nutrientes como para la diversidad de especies nativas en los sistemas terrestres y acuáticos. De hecho, debido a los impactos a largo plazo en las redes alimentarias, los insumos de Nr se consideran ampliamente como el problema de contaminación más crítico en los sistemas marinos. [8] Tanto en los ecosistemas terrestres como en los acuáticos, las respuestas al enriquecimiento de N varían; sin embargo, un tema general recurrente es la importancia de los umbrales (p. ej., saturación de nitrógeno ) en la capacidad de retención de nutrientes del sistema. Para controlar la cascada de N, debe haber integración de disciplinas científicas y más trabajo sobre las tasas de almacenamiento y desnitrificación de Nr . [34]
{{cite book}}: |journal=ignorado ( ayuda )