La queratina del cabello es un tipo de queratina que se encuentra en el cabello y las uñas .
El folículo piloso , que se origina a partir de la epidermis embrionaria, evoluciona hasta convertirse en una de las estructuras más complejas del cuerpo humano, compuesta por 7 u 8 secciones de tejido distintas. [1] La base del folículo piloso contiene el bulbo, que alberga fibroblastos dérmicos conocidos como papila dérmica , cruciales para la morfogénesis y la actividad cíclica del folículo piloso. Rodeando estas células se encuentra la región de células de la matriz, el compartimento proliferativo del folículo piloso, responsable de la formación de diferentes compartimentos del folículo (excepto la SRO) y de la producción de elementos estructurales cruciales del cabello: las queratinas del cabello y las proteínas asociadas conocidas como KAP. [1]
La queratina es una proteína fibrosa crucial que se encuentra en los animales y constituye estructuras resistentes como el pelo, las plumas, las uñas y los cuernos. Se clasifica según el origen del tejido y el contenido de azufre: las queratinas blandas tienen menos azufre, mientras que las queratinas duras, que se encuentran en el pelo y las garras, contienen más azufre, lo que crea una estructura más fuerte. [2] Las queratinas pertenecen a dos tipos: tipo I ácida y tipo II neutra-básica, que se clasifican a su vez en tipo I a y b, y tipo II a y b. El paso inicial en la formación de la queratina es la alineación de los polipéptidos de queratina tipo I y tipo II para crear un heterodímero, que luego se agrega en unidades estructurales de orden superior. [2] De manera similar a otras proteínas de subunidades de filamentos intermedios , existe una estructura secundaria predominante: un dominio alfa-helicoidal central bien conservado compuesto por cuatro segmentos de bobina enrollada junto con dominios terminales no helicoidales que varían en secuencias y longitudes [14]. Recientes hallazgos sugieren que la interacción entre queratinas blandas ácidas y básicas se inicia con la creación de un heterodímero. Este heterodímero comprende una queratina monomérica ácida y una básica. Dos de estos heterodímeros luego se combinan para formar un tetrámero, que posteriormente se polimeriza, lo que da como resultado la formación de la estructura filamentosa final de 10 nanómetros. [3]
Debido a su función como estabilizadores estructurales en las células epiteliales, los filamentos de queratina han despertado un interés significativo en la biología, la embriología, la patología y la dermatología. Esta función fundamental del citoesqueleto se extiende más allá de los niveles de las células individuales. Por lo general, los filamentos de queratina se integran en los desmosomas (ver Fig. 1b, d) y los hemidesmosomas, contribuyendo no solo a la estabilidad entre células sino también a la unión a la membrana basal y al tejido conectivo dentro de un epitelio particular. [4] En los epitelios no estratificados (simples) de los órganos internos que experimentan un estrés mecánico mínimo, solo unos pocos tipos de queratina forman filamentos escasamente distribuidos dentro del citoplasma. Sin embargo, una cantidad más sustancial de tipos de queratina participan en el marco citoesquelético de filamentos intermedios de los epitelios escamosos, que se vuelve más prominente en los epitelios estratificados cornificados como la epidermis que cubre la superficie externa del cuerpo. Aquí, las queratinas son abundantes y densamente empaquetadas, formando tonofilamentos. [4]
Recientemente se ha prestado atención a las notables capacidades de cicatrización de heridas y la excelente biocompatibilidad de la queratina derivada del cabello humano. Si bien las proteínas de queratina recombinante producidas mediante tecnología de ADN recombinante ofrecen una mayor pureza en comparación con la queratina extraída, sus propiedades de cicatrización de heridas no han sido claras. Se expresaron dos queratinas de tricocito recombinantes (queratina capilar humana tipo I 37 y queratina capilar humana tipo II 81) utilizando un sistema de expresión bacteriana y, posteriormente, formando nanopartículas de queratina recombinante (RKNP) mediante dispersión ultrasónica. [5] Se ha revelado que las RKNP impulsaron significativamente la proliferación y migración celular en entornos de laboratorio. Además, cuando se aplicaron a heridas dérmicas in vivo, las RKNP facilitaron una mejor cicatrización de las heridas, lo que llevó a una mejor epitelización, vascularización, deposición de colágeno y remodelación. Es importante destacar que las pruebas de biocompatibilidad in vivo no mostraron signos de toxicidad sistémica. Las RKNP tienen potencial como un enfoque prometedor para avanzar en la cicatrización de heridas y sugieren nuevas vías para desarrollar biomateriales basados en queratina. [5]
Se realizaron estudios de eficacia de hemostasia in vivo utilizando modelos de rata de punción hepática y lesión de la arteria femoral. Para ambos modelos, se aplicaron K37 y K81 (10 mg) para cubrir las áreas de la herida. En el modelo de punción hepática, el tiempo de sangrado disminuyó significativamente con K37 recombinante (aproximadamente 38 s) y K81 (aproximadamente 40 s) en comparación con el vehículo solo (aproximadamente 170 s, p < .01), con una pérdida total de sangre notablemente reducida (p < .01). [6] Además, en el modelo de lesión de la arteria femoral, las proteínas de queratina recombinante redujeron significativamente el tiempo de sangrado en comparación con el grupo de control (aproximadamente 50 s frente a 270 s). En particular, K37 y K81 exhibieron efectos hemostáticos más fuertes que las queratinas extraídas (aproximadamente 80 s) en el tratamiento de la lesión hepática de rata. Además, las proteínas de queratina recombinantes demostraron una sólida capacidad para promover la formación de un coágulo de fibrina en el lugar de la lesión, deteniendo eficazmente el sangrado. En consecuencia, las queratinas de cabello humano recombinantes ofrecen potencial para desarrollar nuevos productos hemostáticos basados en biomateriales de queratina. [6]
Existen dos tipos de queratina capilar:
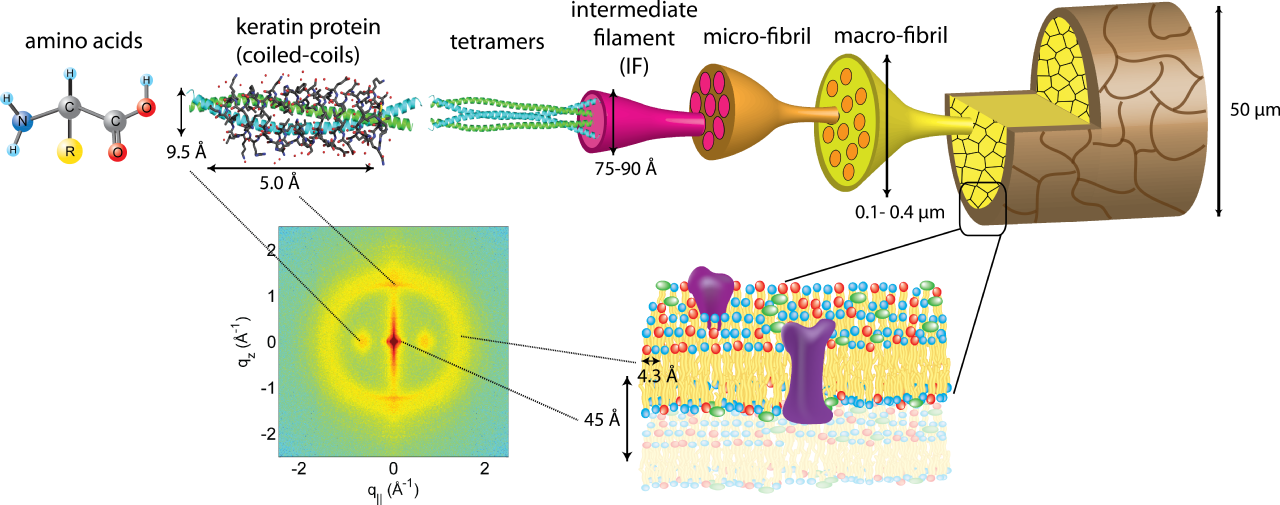
El tallo del cabello está compuesto principalmente de queratinas capilares y sus proteínas asociadas (KRTAP). Las KRTAP son productos de diversas familias de genes resultantes de eventos de duplicación genética en su historia evolutiva. Estos genes son típicamente pequeños, y comprenden un solo exón de menos de 1000 pares de bases de longitud. Durante la última década, se han identificado numerosos genes KRTAP en mamíferos, incluidos los humanos. Se clasifican en tres grupos según su composición de aminoácidos: alto contenido de azufre (con <30 mol % de cisteína), ultraalto contenido de azufre (>30 mol % de cisteína) y alto contenido de glicina/tirosina. [7] Las queratinas capilares forman filamentos intermedios (KIF) dentro de los tricocitos, células especializadas que contribuyen a la formación del cabello. A medida que estas células se mueven hacia arriba en la corteza, los KIF se agregan, rodeados por un espacio llamado matriz. Los KRTAP, también conocidos como KAP, son una parte importante de esta matriz entre los KIF. Se sugiere que los KRTAP desempeñan un papel en el establecimiento de una red reticulada con los KIF, contribuyendo a la creación del tallo rígido del cabello. [7]
Durante el crecimiento del pelo, a medida que las células del bulbo folicular se transforman rápidamente en queratinocitos pilosos corticales o cuticulares, se activan aproximadamente entre 50 y 100 genes de queratina a nivel transcripcional. [8] Sin embargo, este intrincado proceso se puede simplificar en unas pocas familias de genes altamente conservadas. En los queratinocitos corticales, son evidentes patrones distintos de expresión de genes de queratina, lo que indica la presencia de diferentes procesos de transcripción jerárquica entre varios tipos de células. El examen de las regiones promotoras de genes de queratina revela motivos de secuencia conservados que podrían gobernar estos rasgos específicos de las células. [8] Además, a través del aislamiento de genes de queratina cuticular de ovejas y humanos relacionados, se han descubierto motivos de ADN conservados y patrones de expresión durante la diferenciación de las células de la cutícula. Además, la expresión de IF de folículos de lana de oveja y genes de queratina con alto contenido de azufre en ratones transgénicos sugiere que los elementos reguladores del ADN y las proteínas asociadas con los genes de queratina pilosa mantienen una conservación funcional en todas las especies de mamíferos. [8]
La queratina constituye una gran familia multigénica conocida como citoqueratinas. Estas citoqueratinas se expresan de forma diferencial en varios tipos epiteliales y se han estudiado ampliamente como marcadores del cáncer de mama. Se clasifican en citoqueratinas de tipo I ácidas y de tipo II básicas a neutras. [9] La red de filamentos intermedios está formada por el emparejamiento necesario de cantidades iguales de queratinas de tipo I y tipo II. Si bien las queratinas del cabello, como KRT81, son típicas de estructuras queratinizadas duras como el cabello y las uñas, se cree que sirven como proteínas estructurales específicas de estos órganos sin expresión en otras partes, como la glándula mamaria.
KRT81, una queratina capilar de tipo II, es una proteína capilar importante que se expresa en la corteza capilar. Curiosamente, a pesar de estar asociada normalmente a las estructuras capilares, se ha observado la expresión de KRT81 en la línea celular de cáncer de mama humano SKBR3 y en los ganglios linfáticos metastásicos de carcinomas de mama, pero no en las células epiteliales mamarias normales. Además, se descubrió que la KRT81 expresada era una isoforma truncada en 5′ (ΔHb1), y que la proteína de longitud completa no se expresaba. [9] Sin embargo, la función exacta de esta forma truncada en las células de cáncer de mama sigue sin estar clara.
El análisis Western blot detectó la presencia del KRT81 de 55 kDa completo en varias líneas celulares de cáncer de mama humano (MCF7, SKBR3, MDA-MB-231), células epiteliales mamarias humanas normales (HMEC) y células no neoplásicas (MCF10A). [9] La reacción en cadena de la polimerasa con transcripción inversa confirmó la expresión del KRT81 de longitud completa, que abarca su región 5', en células mamarias. Los exámenes inmunohistoquímicos y de inmunofluorescencia localizaron el KRT81 dentro del citoplasma. Además, en las células MDA-MB231 con KRT81 inactivado, la zimografía ilustró una actividad reducida de MMP9, mientras que los ensayos de raspado e invasión demostraron una disminución de la migración celular y de las capacidades de invasión. [9] Esto presenta la primera evidencia de la expresión completa de KRT81 tanto en células epiteliales mamarias normales como en células de cáncer de mama. Además, los hallazgos sugieren que el KRT81 desempeña un papel en la migración e invasión de células de cáncer de mama.