
Las opsinas visuales de vertebrados son una subclase de las opsinas ciliares y median la visión en los vertebrados. Incluyen las opsinas de los conos y bastones humanos . A menudo se las abrevia como opsinas , ya que fueron las primeras opsinas descubiertas y siguen siendo las opsinas más estudiadas. [1]
La opsina se refiere estrictamente a la apoproteína (sin retinal unido). Cuando una opsina se une al retinal para formar una holoproteína , se la denomina proteína retinilideno . Sin embargo, a menudo se ignora la distinción y la opsina puede referirse vagamente a ambas (independientemente de si el retinal está unido o no).
Las opsinas son receptores acoplados a proteína G (GPCR) y deben unirse al retinal —normalmente 11- cis -retinal — para ser fotosensibles, ya que el retinal actúa como cromóforo . Cuando la proteína retinilideno absorbe un fotón, el retinal se isomeriza y es liberado por la opsina. El proceso que sigue a la isomerización y renovación del retinal se conoce como ciclo visual . El 11- cis -retinal libre es fotosensible y tiene su propia sensibilidad espectral de 380 nm. [2] Sin embargo, para desencadenar la cascada de fototransducción , el proceso que subyace a la señal visual, el retinal debe estar unido a una opsina cuando se isomeriza. La proteína retinilideno tiene una sensibilidad espectral que difiere de la del retinal libre y depende de la secuencia de la opsina.
Si bien las opsinas solo pueden unirse al retinal, existen dos formas de retinal que pueden actuar como cromóforo para las opsinas visuales de los vertebrados:
Los animales que viven en tierra y los peces marinos forman sus pigmentos visuales exclusivamente con retinal 1. Sin embargo, muchos peces de agua dulce y anfibios también pueden formar pigmentos visuales con retinal 2, dependiendo de la activación de la enzima retinal-3,4-desaturasa (GO:0061899). Muchas de estas especies pueden cambiar entre estos cromóforos durante su ciclo de vida, para adaptarse a un hábitat cambiante. [3] [4]
La isomerización del 11- cis -retinal en todo- trans -retinal por la luz induce un cambio conformacional en la proteína que activa la vía de fototransducción .
Hay dos clases de opsina visual en vertebrados, que se diferencian según se expresen en fotorreceptores de bastones o de conos.
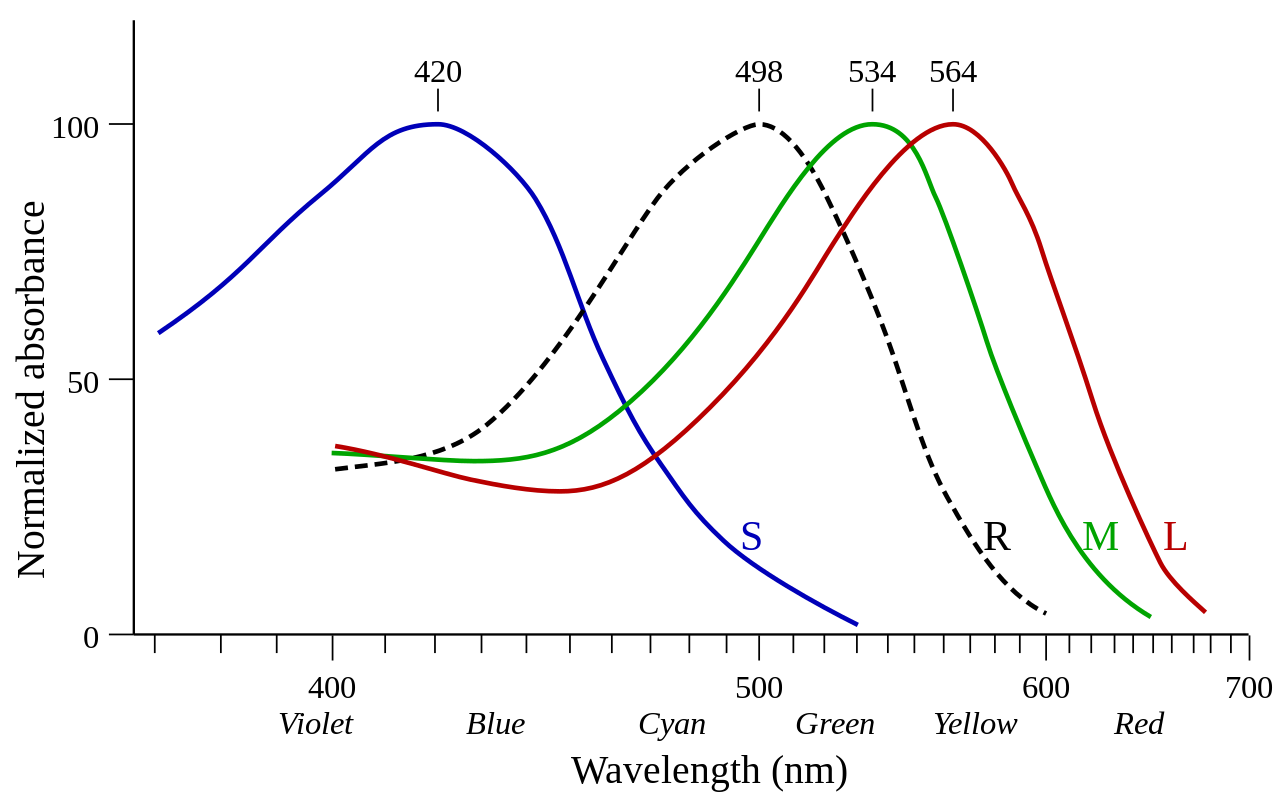
Las opsinas expresadas en las células de los conos se denominan opsinas de cono. [1] Las opsinas de cono se denominan fotopsinas cuando no están unidas al retinal y yodopsinas cuando están unidas al retinal. [1] Las opsinas de cono median la visión fotópica (luz del día). Las opsinas de cono se subdividen según la sensibilidad espectral de su yodopsina, es decir, la longitud de onda en la que se observa la mayor absorción de luz ( λ max ). [7]
Las opsinas expresadas en las células de los bastones se denominan opsinas de bastones. Las opsinas de bastones se denominan escotopsinas cuando no están unidas al retinal y rodopsinas o porfiropsinas cuando están unidas al retinal (1 y 2, respectivamente). Las opsinas de bastones median la visión escotópica (luz tenue). [8] En comparación con las opsinas de los conos, la sensibilidad espectral de la rodopsina es bastante estable y no se desvía mucho de los 500 nm en ningún vertebrado.
Los vertebrados actuales suelen tener cuatro clases de opsinas de cono (LWS, SWS1, SWS2 y Rh2), así como una clase de opsinas de bastón (rodopsina, Rh1), todas las cuales fueron heredadas de ancestros vertebrados tempranos. Estas cinco clases de opsinas visuales de vertebrados surgieron a través de una serie de duplicaciones genéticas comenzando con LWS y terminando con Rh1, según el cladograma de la derecha; esto sirve como un ejemplo de neofuncionalización . Desde entonces, cada clase ha evolucionado en numerosas variantes. [9] [10] Las relaciones evolutivas, deducidas utilizando la secuencia de aminoácidos de las opsinas, se utilizan con frecuencia para categorizar las opsinas de cono en su clase respectiva. [1] Los mamíferos perdieron las clases Rh2 y SWS2 durante el cuello de botella nocturno . Los ancestros de los primates desarrollaron más tarde dos opsinas LWS (LWS y MWS), dejando a los humanos con 4 opsinas visuales en 3 clases.
George Wald recibió el Premio Nobel de Fisiología o Medicina en 1967 por sus experimentos en la década de 1950 que demostraron la diferencia en la absorbancia de estas fotopsinas (ver imagen). [11]
Como lo deja claro la Fig. 11a, la MSP es de poca utilidad para definir las sensibilidades espectrales de los conos, excepto cerca del fotopigmento λmax. Las grandes discrepancias entre la MSP y otras estimaciones de las sensibilidades espectrales de los conos surgen debido a la pequeña relación señal/ruido de las mediciones de la MSP.