Los dictiostélidos ( Dictyostelia/Dictyostelea , ICZN , Dictyosteliomycetes , ICBN ) o mohos mucilaginosos celulares son un grupo de mohos mucilaginosos o amebas sociales .

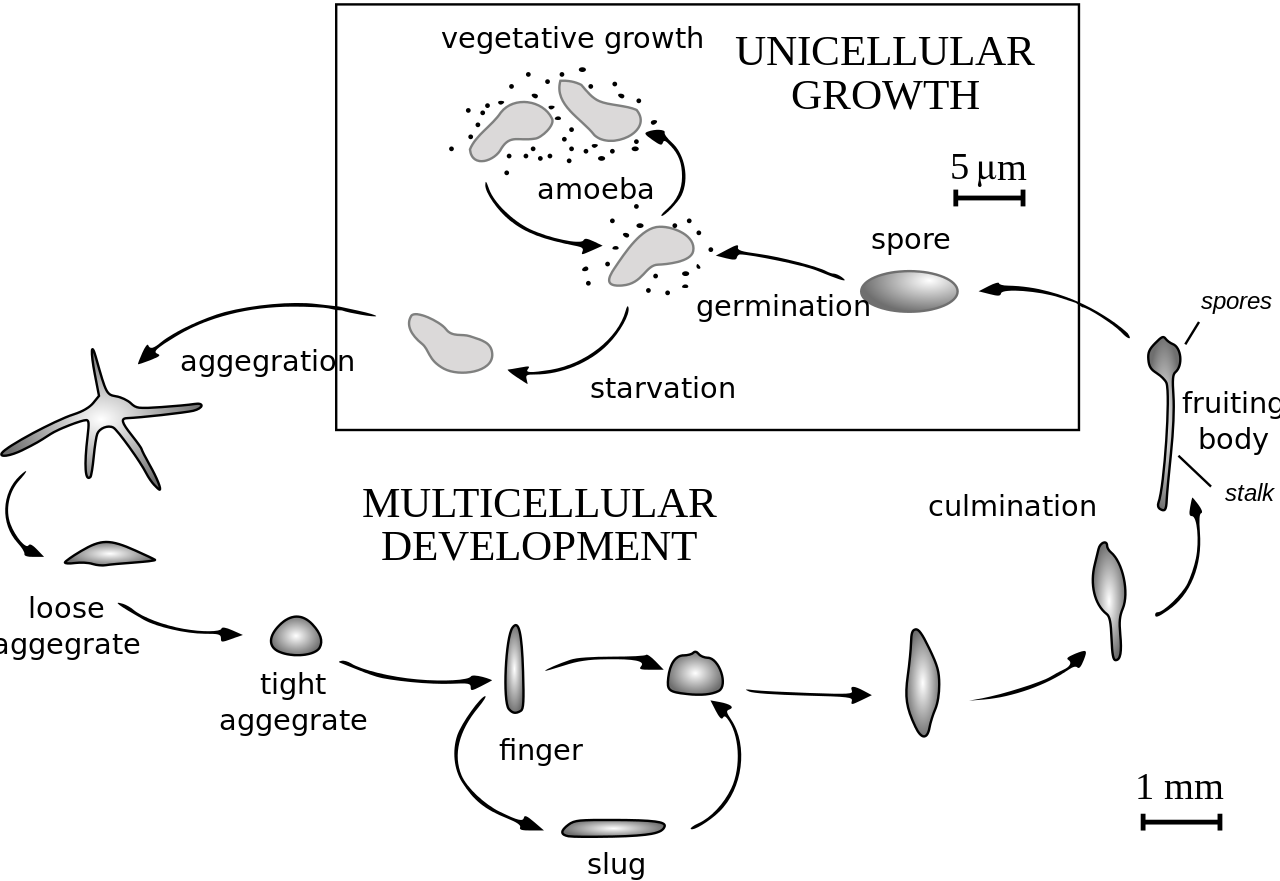
Cuando el alimento (normalmente bacterias) está fácilmente disponible, los dictiostélidos se comportan como amebas individuales, que se alimentan y se dividen normalmente. Sin embargo, cuando el suministro de alimento se agota, se agregan para formar un conjunto multicelular, llamado pseudoplasmodio, grex o babosa (que no debe confundirse con el molusco gasterópodo llamado babosa ). El grex tiene una parte anterior y posterior definidas, responde a los gradientes de luz y temperatura y tiene la capacidad de migrar. En las circunstancias adecuadas, el grex madura formando un sorocarpo (cuerpo fructífero) con un tallo que sostiene uno o más soros (bolas de esporas). Estas esporas son células inactivas protegidas por paredes celulares resistentes y se convierten en nuevas amebas una vez que el alimento está disponible.
En Acytostelium , el sorocarpio está sostenido por un tallo compuesto de celulosa , pero en otros dictiostélidos el tallo está compuesto de células, que a veces ocupan la mayoría de las amebas originales. Con unas pocas excepciones, estas células mueren durante la formación del tallo, y existe una correspondencia definida entre partes del grex y partes del cuerpo fructífero. La agregación de amebas generalmente tiene lugar en corrientes convergentes. Las amebas se mueven utilizando pseudópodos filosos y son atraídas por sustancias químicas producidas por otras amebas. En Dictyostelium discoideum , la agregación está señalizada por AMPc , pero otros utilizan sustancias químicas diferentes. En la especie Dictyostelium purpureum , la agrupación es por parentesco, no solo por proximidad.
Dictyostelium se ha utilizado como organismo modelo en biología molecular y genética , y se estudia como un ejemplo de comunicación celular , diferenciación y muerte celular programada . También es un ejemplo interesante de la evolución de la cooperación y el engaño. [1] [2] [3] Una gran cantidad de datos de investigación sobre D. discoideum está disponible en línea en DictyBase .

El mecanismo que subyace a la agregación de las amebas se basa en el monofosfato de adenosina cíclico (AMPc) como molécula señal. Una célula, la fundadora de la colonia, comienza a secretar AMPc en respuesta al estrés. Otras detectan esta señal y responden de dos maneras:
El efecto de esto es transmitir la señal a toda la población cercana de amebas y provocar un movimiento hacia el área de mayor concentración de AMPc.
Dentro de una célula individual, el mecanismo es el siguiente:
Debido a que la concentración interna de AMPc inactiva el receptor de AMPc externo, cada célula muestra un comportamiento oscilatorio . Este comportamiento produce hermosas espirales que se observan en colonias convergentes y recuerdan a la reacción de Belousov-Zhabotinsky y a los autómatas celulares cíclicos bidimensionales .
El genoma completo de Dictyostelium discoideum fue publicado en Nature en 2005 por el genetista Ludwig Eichinger y colaboradores. [4] El genoma haploide contiene aproximadamente 12.500 genes en 6 cromosomas. A modo de comparación, el genoma humano diploide tiene entre 20.000 y 25.000 genes (representados dos veces) en 23 pares de cromosomas. Hay un alto nivel de los nucleótidos adenosina y timidina (~77 %), lo que conduce a un uso de codones que favorece más adenosinas y timidinas en la tercera posición. Las repeticiones en tándem de trinucleótidos son abundantes en Dictyostelium, que en los humanos causa trastornos de repetición de trinucleótidos .
El desarrollo sexual puede ocurrir cuando las células ameboides están privadas de su suministro de alimento bacteriano y existen condiciones húmedas y oscuras. [5] Tanto las cepas heterotálicas como las homotálicas de Dictostelium pueden experimentar apareamiento. El desarrollo sexual heterotálico ha sido estudiado más extensamente en D. discoideum , y el desarrollo sexual homotálico ha sido estudiado más a fondo en D. mucoroides . [6] Los apareamientos heterotálicos se inician por la fusión de células haploides (gametos) de dos cepas de tipo de apareamiento opuesto. Esto contrasta con las cepas homotálicas que parecen expresar ambos tipos de apareamiento. [7]
El apareamiento se inicia mediante la gametogénesis, que produce gametos pequeños y móviles que se fusionan para formar una pequeña célula binucleada . El volumen de la célula binucleada aumenta luego para producir una célula binuclear gigante. A medida que avanza el crecimiento, los núcleos se hinchan y luego se fusionan formando una verdadera célula gigante cigoto diploide. Mientras esto ocurre, las amebas han estado experimentando quimiotaxis inducida por AMPc hacia la superficie de la célula gigante. Esto forma un agregado celular y en el centro del agregado la célula gigante cigoto ingiere las amebas circundantes. La fagocitosis es seguida por la digestión de las amebas ingeridas. A continuación, el cigoto forma un macroquiste caracterizado por una vaina de celulosa extracelular circundante. Después de que se forma el macroquiste, normalmente permanece inactivo durante un período antes de que pueda ocurrir la germinación. [8] Dentro del macroquiste, el cigoto diploide experimenta meiosis seguida de divisiones mitóticas sucesivas. Cuando el macroquiste germina libera muchas células ameboides haploides.
El árbol filogenético de Dictyostelium ha sufrido múltiples cambios en las últimas décadas. El primer dictiostélido que se describió fue Dictyostelium mucoroides en 1869 por Osker Brefeld, y el descubrimiento original de Dictyostelium discoideum ocurrió en 1935, [9] con descubrimientos adicionales de Kenneth Raper , seguidos por esfuerzos globales liderados por James Cavender y colaboradores. Dictyostelium discoideum fue clasificado inicialmente bajo "hongos inferiores", pero la clasificación ha cambiado desde entonces a bajo el filo Amoebozoa donde se encuentra actualmente. [4]
Las agrupaciones dentro del árbol filogenético de los dictiostélidos han sufrido frecuentes reordenamientos debido a la disponibilidad de nuevas evidencias. La mayoría de las filogenias de los dictiostélidos aceptadas actualmente utilizan la secuenciación genómica y el ADN ribosómico de subunidades pequeñas (ssu-rDNA). Los dictiostélidos pueden subdividirse en cuatro grupos. En particular, el grupo 4 contiene la especie Dictyostelium discoideum y se diferencia de los otros grupos por el uso de AMPc como un atrayente emitido durante la agregación. [10]
Las calibraciones fósiles indican que la clase de los dictiostélidos originalmente divergió en dos ramas principales hace aproximadamente 520 millones de años. Las teorías actuales especulan que la formación de tallos y esporas de los dictiostélidos originalmente evolucionó como una adaptación a las formaciones glaciares globales. Es probable que la subdivisión posterior de las especies de dictiostélidos surgiera a medida que la mayoría de las formaciones glaciares se derretían. La mayoría de las especies de los grupos principales 1, 2 y 3 muestran una capacidad de enquistamiento que les permite sobrevivir a bajas temperaturas, pero las esporas han mostrado una mayor capacidad para resistir temperaturas más bajas. El grupo 4 se diferencia de otros grupos principales en una falta general de capacidad para enquistarse, pero sus esporas han mostrado una mejor resistencia a temperaturas más bajas en relación con las esporas de otros grupos. [11]
La filogenia interna de los Dictyostelids se muestra en el cladograma . [12]
Clase Dictyostelia Lister 1909 en Olive 1970 [13]
Dictyostelium comparte muchas características moleculares con los macrófagos , el huésped humano de Legionella . La composición del citoesqueleto de D. discoideum es similar a la de las células de los mamíferos, al igual que los procesos impulsados por estos componentes, como la fagocitosis, el tráfico de membranas, el tránsito endocítico y la clasificación de vesículas. Al igual que los leucocitos, D. discoideum posee capacidad quimiotáctica. Por lo tanto, D. discoideum representa un sistema modelo adecuado para determinar la influencia de una variedad de factores de la célula huésped durante las infecciones por Legionella . [14]