Los ribosomas contienen dos principales tipos de ARNr que forman dos subunidades: la subunidad mayor (LSU, Large Sub Unit por sus siglas en inglés, que es una ribozima que cataliza la formación de enlaces peptídicos), y la subunidad menor (SSU, Small Sub Unit).Tradicionalmente, los ARN ribosómicos se denominan según su coeficiente de sedimentación, medido en unidades Svedberg (S).Las arqueas contienen un operón de rDNA o bien múltiples copias.Debido a su estructura especial y a su patrón de transcripción, los grupos de genes de ARNr se suelen denominar «ADN ribosomal (ADNr)» (nótese que el término no se refiere a que los ribosomas contengan ADN).Las características del ARN ribosómico son importantes en evolución, taxonomía y medicina.
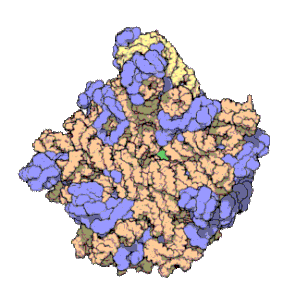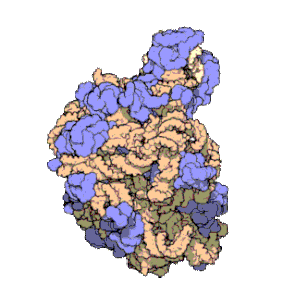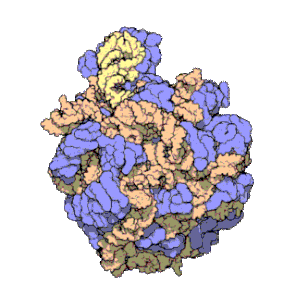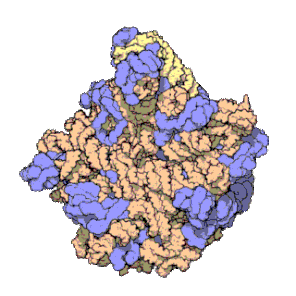
Estructura 3D de la subunidad 50S de
Haloarcula marismortui
. Las proteínas se muestran en verde, el ARNr 23S en naranja y el ARNr 5S en amarillo.
[
1
]