
La selección sexual en los mamíferos es un proceso cuyo estudio comenzó con las observaciones de Charles Darwin sobre la selección sexual , incluida la selección sexual en humanos y en otros mamíferos , [1] que consiste en la competencia entre machos y la elección de pareja que moldean el desarrollo de futuros fenotipos en una población para una especie determinada . [2] [3]

Un buen ejemplo de selección intrasexual, en la que los machos luchan por el dominio sobre un harén de hembras, es el elefante marino , un gran mamífero oceánico del género Mirounga . Hay dos especies: el elefante marino del norte ( M. angustirostris ) y el elefante marino del sur ( M. leonina ), el carnívoro más grande que vive en la actualidad. Ambas especies muestran un dimorfismo sexual extremo , posiblemente el más grande de cualquier mamífero, con los machos del sur típicamente cinco a seis veces más pesados que las hembras. [4] Mientras que las hembras promedian de 400 a 900 kilogramos (880 a 1,980 libras) y 2,6 a 3 metros (8,5 a 9,8 pies) de largo, los machos promedian de 2,200 a 4,000 kilogramos (4,900 a 8,800 libras) y 4,2 a 5 metros (14 a 16 pies) de largo. [5] [6] El toro de tamaño récord, abatido a tiros en Possession Bay, Georgia del Sur , el 28 de febrero de 1913, medía 6,85 metros (22,5 pies) de largo y se estimó que pesaba 5000 kilogramos (11 000 libras). [7] [8] El peso máximo de una hembra es de 1000 kilogramos (2200 libras) con una longitud de 3,7 metros (12 pies).
Los machos llegan a las colonias antes que las hembras y luchan por el control de los harenes. [9] El gran tamaño corporal confiere ventajas en la lucha. El comportamiento agonístico de los machos da lugar a una jerarquía de dominancia, y el acceso a los harenes y la actividad reproductiva se determinan por el rango. [10] Los machos dominantes o "amos del harén" establecen harenes de varias docenas de hembras. Los machos menos exitosos no tienen harenes, pero pueden intentar copular con las hembras de un macho de harén cuando el macho dominante no está mirando. Un macho dominante debe permanecer en su territorio para defenderlo, lo que puede significar meses sin comer, viviendo de su reserva de grasa. Algunos machos han permanecido en tierra durante más de tres meses sin comida. Dos machos que pelean usan su peso y sus colmillos uno contra el otro. El resultado rara vez es fatal, y el macho derrotado huye; sin embargo, los machos sufren desgarros y cortes severos. Los machos comúnmente vocalizan con un rugido de tos que sirve tanto para el reconocimiento individual como para la evaluación del tamaño. Los conflictos entre machos de alto rango se resuelven más a menudo con posturas y vocalizaciones que con contacto físico. [10]

En el caso de la selección intrasexual , los machos adornados pueden obtener una ventaja reproductiva sin la intervención de la preferencia femenina. Esta ventaja será conferida por las armas utilizadas en el proceso de resolución de disputas, como las que se refieren a los derechos territoriales. El uso de la ornamentación sexual como un dispositivo de señalización para crear una jerarquía de dominio entre los machos, también conocida como orden jerárquico , permite que la lucha se lleve a cabo sin lesiones o fatalidades excesivas. Es predominantemente cuando dos machos opuestos están tan estrechamente emparejados, como se encontraría en los machos que no se han establecido en una jerarquía de dominio, que no se pueden encontrar asimetrías y la confrontación se intensifica hasta un punto en que las asimetrías deben probarse mediante el uso agresivo de la ornamentación.
La frecuencia con la que los hombres interactúan físicamente entre sí, y de qué manera, se puede entender mejor aplicando la teoría de juegos desarrollada para la biología, sobre todo por John Maynard Smith . [11]

Los orígenes evolutivos del largo cuello de la jirafa ( Giraffa camelopardalis ) son controvertidos. La "hipótesis del ramoneador competidor", propuesta originalmente por Charles Darwin y aceptada durante mucho tiempo , ha sido puesta en tela de juicio. Originalmente, los científicos creían que el alargamiento del cuello de la jirafa había sido el resultado de la selección natural que actuaba en relación con el comportamiento de búsqueda de alimento , donde se suponía que los cuellos más largos permitían a los individuos favorecidos recolectar alimentos inaccesibles para otros animales. [12] Pero a pesar de que la altura total de la jirafa es de unos 6 metros, todavía suele alimentarse a unos 2 metros sobre el suelo. [13] Además, los competidores de la jirafa, el kudú, el impala y el steenbok, no se alimentan por encima de los 2 metros y prefieren hacerlo también a la altura de los hombros, en lugar de a la altura máxima que podrían alcanzar. [14]
Una explicación alternativa para el origen de los cuellos largos en las jirafas es la selección sexual. Las jirafas macho a menudo se besan con otros machos para mostrar dominio. [15] Hay seis criterios que deben cumplirse para que el cuello exagerado se clasifique como resultado de la selección sexual. La característica debe ser más exagerada en uno de los sexos; debe usarse para indicar dominio; no tener beneficios directos para la supervivencia; costarle al organismo en términos de supervivencia u otros factores (por ejemplo, energética [16] [17] ); debe observarse una alometría positiva. [18] Pero la historia evolutiva muestra que el aumento de la longitud del cuello no está correlacionado con aumentos en otras partes del cuerpo, lo que se esperaría de la selección de forrajeo, por lo que la selección sexual puede ser una explicación más satisfactoria. [19] Los estudios no han logrado resolver las causas involucradas: tal vez el cuello fue el resultado de ambas u otras fuerzas. [20]
Los mecanismos precopulatorios determinan quién engendrará una descendencia antes de la relación sexual. La competencia entre machos es el mayor mecanismo precopulatorio en los mamíferos. El dimorfismo sexual es un resultado de la competencia entre machos que se observa fácilmente en las especies.


La competencia entre machos para copular con el sexo opuesto es algo que se observa a menudo en los mamíferos . Los elefantes africanos promueven fuertemente la competencia entre machos. Los elefantes crecen continuamente a lo largo de su vida. A medida que los machos envejecen, también experimentan lapsos cada vez mayores de celo , una excitación sexual violenta, y la mayor parte del éxito reproductivo se da en los machos en celo, ya que les ayuda a ganar peleas. [21] Una pelea entre un macho en celo y uno que no lo está puede resultar en la muerte de este último.
Se sabe que las especies con una intensa competencia entre machos presentan el mayor dimorfismo de tamaño. [22] Por ejemplo, las hembras de oso negro americano ( Ursus americanus ) son entre un 20 y un 40 % más pequeñas que los machos. [23]
Los mamíferos machos también pueden competir por harenes , al igual que los elefantes marinos que compiten ferozmente por ellos. A medida que los mamíferos alcanzan la madurez sexual, surgen las características sexuales secundarias . [24] Los elefantes marinos tienen una probóscide en el macho adulto, que se utiliza para proyectar ruidos fuertes, que se escuchan con frecuencia durante la temporada de apareamiento. [25] Los elefantes marinos con una probóscide más grande emiten sonidos más bajos que los machos con una probóscide más pequeña y son los machos más grandes de una colonia. La protección de la pareja es un factor importante en la competencia entre machos para asegurar la fertilización de una descendencia y, cuando tiene éxito, ayuda a supervisar y cortejar a la hembra. [26] Especialmente previene la competencia de espermatozoides , ya que asegura el éxito reproductivo. Este proceso puede activarse cuando se envía una señal poscoital a un macho para que la hembra mantenga la vigilancia. [27] Los tapones de apareamiento son una forma de protección de la pareja que ha demostrado mostrar la elección femenina precopulatoria. [28] Los tapones copuladores son comúnmente acelulares y se cree que están hechos de proteínas de las vesículas seminales. [29] El ADN extraído de los tapones copulatorios muestra que las hembras evitan aparearse con parientes cercanos.

Durante la temporada de cría , los mamíferos llaman al sexo opuesto. Los koalas machos que son más grandes emitirán un sonido diferente al de los koalas más pequeños. Los machos más grandes que se buscan rutinariamente se llaman sementales. Las hembras eligen a los sementales debido a los beneficios indirectos que su descendencia podría heredar, como cuerpos más grandes. [30] Los no sementales y las hembras no varían en su masa corporal y pueden rechazar a un macho gritando o golpeándolo. La competencia macho-macho rara vez se exhibe en los koalas. [31] La señalización acústica es un tipo de llamada que se puede usar desde una distancia significativa que codifica la ubicación, la condición y la identidad de un organismo. [32] Los murciélagos de alas en saco muestran señalización acústica, que a menudo se interpreta como canciones. Cuando las hembras escuchan estas canciones, llamadas "silbidos", llaman a los machos para que se reproduzcan con un chillido propio. Esta acción se denomina "llamado de los sexos". [33] Los ciervos rojos y las hienas manchadas , junto con otros mamíferos, también realizan señalización acústica. [34] [35]

La testosterona es un factor determinante para lograr el éxito de la fecundación. Los cuernos de los carneros de las Montañas Rocosas son curvados, grandes en comparación con los pequeños cuernos de las hembras. Cuanto más grandes son los cuernos, más testosterona hay en el macho. Esto es importante porque el rango social tiene una correlación positiva con la longitud de los cuernos. [24] El rango social hace que un grupo de hembras tienda a copular. [36] La testosterona también aparece más en las especies polígamas que en las monógamas. [37]
Las hembras poliándricas tienen dos o más parejas de apareamiento durante el celo. Las hembras tienen más probabilidades de encontrar una nueva pareja cuando su macho actual tuvo un gran número de paternidades el año anterior o su macho actual era mayor. Se presume que esto tiene un efecto en la descendencia y les otorga una mayor diversidad genética. [38] [39]
_Tswalu.jpg/1280px-Meerkat_(Suricata_suricatta)_Tswalu.jpg)
La inversión de roles sexuales es el cambio en el comportamiento de una especie específica con respecto a su patrón ordinario. La inversión de roles sexuales apoya muy fuertemente el dimorfismo sexual. [40] La competencia entre hembras es una anormalidad común dentro de los animales con roles sexuales aceptados. Las hembras invierten en elegir la mejor pareja posible porque tienen una mayor participación en la crianza de sus crías que los machos (gestación y lactancia). La gestación y la lactancia consumen energía, lo que significa que su competencia por los recursos es alta. [41] Se observa que las competencias entre hembras tienen como objetivo obtener mejores parejas. Las hembras de suricata adquieren el estatus dominante porque los recursos para la reproducción femenina son escasos. [42] Las hembras dominantes en esta especie son más pesadas y ganan en las competencias contra otras hembras.
La copulación con el sexo opuesto no garantiza la fecundación del óvulo. Los mecanismos poscopulatorios incluyen la competencia espermática [43] [44] [45] y la elección críptica de la hembra . [46] [47]
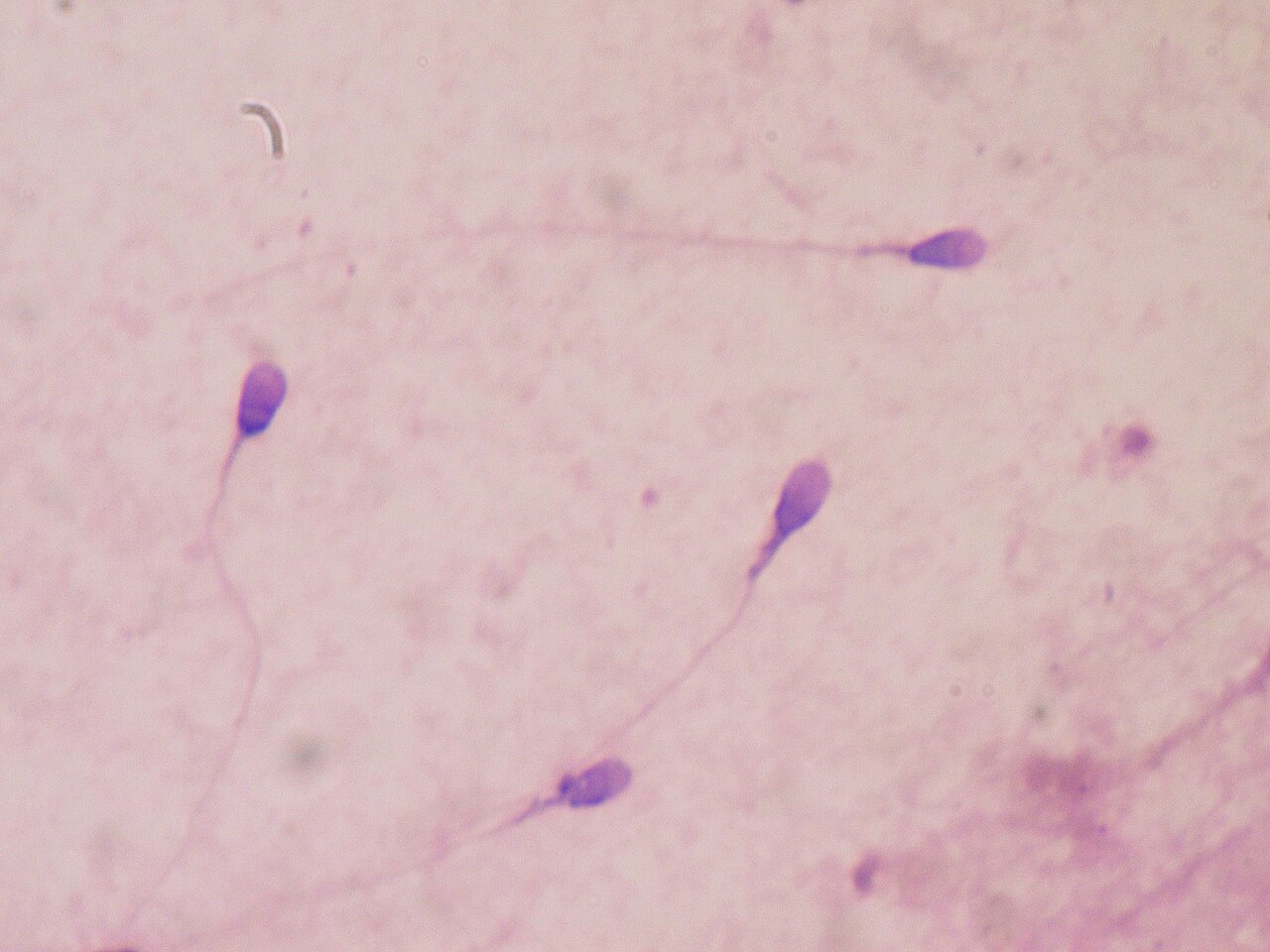
La competencia entre espermatozoides implica que los gametos masculinos intenten fecundar los óvulos primero. Como resultado de la competencia entre espermatozoides, algunos machos de una especie determinada pueden desarrollar testículos y vesículas seminales más grandes. [48] También se observan áreas más grandes de la parte media del esperma que contienen mitocondrias. [37] Se observan testículos y partes medias más grandes en los espermatozoides en los machos que se aparean con múltiples parejas. Una hembra que ha estado con múltiples parejas probablemente dará a luz una descendencia engendrada por el macho que produjo más espermatozoides o que los produjo más rápido. [48] Se descubrió que los primates y roedores con flagelos más largos engendraron más descendencia. [49] La longitud del báculo también está influenciada por la competencia entre espermatozoides en algunas especies de mamíferos. [50]
La elección femenina críptica es un mecanismo poscopulatorio que no se puede observar porque tiene lugar dentro del cuerpo de la hembra. [51] Permite a la hembra tener cierto control sobre quién es el padre de su hijo incluso después de la fertilización. [52]
En algunas especies, las hembras pueden optar por aparearse con más de un macho para evitar el infanticidio o el acoso. El infanticidio se puede prevenir confundiendo a los machos de una colonia determinada. Si la hembra se aparea con varios machos, los machos no sabrán con seguridad quién es el padre de la descendencia. El infanticidio también se puede prevenir eligiendo un macho que la proteja a ella y a la descendencia. [48] El acoso sexual se puede evitar si las hembras se rinden ante los machos y copulan cuando les place. [53]
{{cite journal}}: CS1 maint: varios nombres: lista de autores ( enlace )