[cita requerida] Presentan un caparazón con dos valvas laterales, que se cierran por acción de uno o dos músculos aductores.[cita requerida] Entre los moluscos bivalvos más conocidos podemos nombrar: ostra, almeja, navaja, mejillón, broma de los barcos, coquina, etc.En el borde anterior del manto se distinguen tres pliegues: interno, medio y externo.El pliegue interno es muscular, el medio destaca por su función sensorial y el externo está relacionado con la secreción de la concha.Las valvas también están unidas dorsalmente por el ligamento bisagra, que es una prolongación del periostraco.En todos los bivalvos, salvo en los más primitivos, hay dos ganglios cerebropleurales a cada lado del esófago.[11][12] Los órganos sensoriales de los bivalvos están situados en su mayor parte en los márgenes posteriores del manto.El osfradio es un parche de células sensoriales situado bajo el músculo aductor posterior que puede servir para degustar el agua o medir su turbidez.[12] En el orden Anomalodesmata, el sifón inhalante está rodeado de tentáculos sensibles a la vibración para detectar presas.El corazón tiene tres cámaras: dos aurículas que reciben la sangre de las branquias y un ventrículo único.Algunos bivalvos tienen una sola aorta, pero la mayoría también tienen una segunda, normalmente más pequeña, que sirve a las partes posteriores del animal.[17] Las branquias emparejadas están situadas en la parte posterior y consisten en filamentos tubulares huecos con paredes finas para el intercambio de gases.[18][19] Las ostras, incluida la ostra del Pacífico (Magallana gigas), tienen respuestas metabólicas variables al estrés ambiental, observándose con frecuencia cambios en la tasa de respiración.Éstos clasifican las partículas, rechazando las inadecuadas o demasiado grandes para digerirlas y transportando las demás a la boca.[21] En los bivalvos más avanzados, el agua se introduce en la concha desde la superficie ventral posterior del animal, pasa hacia arriba a través de las branquias y se duplica para ser expulsada justo por encima de la entrada.La estructura de las branquias varía considerablemente y puede servir para clasificar a los bivalvos en grupos.Para evitar ser arrastrado, el pepino de mar se sujeta con hilos bisoños a la garganta del huésped y permanece sin daños.También tienen glándulas pericárdicas que recubren las aurículas del corazón o se adhieren al pericardio, y sirven como órganos de filtración adicionales.Las gónadas se abren en los nefridios o a través de un poro separado en una cámara sobre las branquias.Otras desovan en masa y liberan sus gametos por lotes o todos a la vez.El coste reproductivo de producir estos huevos ricos en energía es elevado y suelen ser más pequeños en número.Por ejemplo, la tellina del Báltico (Macoma balthica) produce pocos huevos de alto valor energético.Los bivalvia son moluscos esencialmente acuáticos, en su mayoría marinos, que habitan con preferencia la región bentónica sublitoral, aunque también se los encuentra hasta las profundidades abisales.Tienen gran valor en los análisis paleoambientales debido a su estrecha relación con el tipo de sustrato, la buena presencia en el registro fósil desde el Cámbrico, su gran diversidad y abundancia en variados paleoambientes, y excelente representación en los medios acuáticos actuales.Muchos bivalvos (especialmente los infaunales) viven con su comisura orientada en posición perpendicualr a la interfase entre el sustrato y el agua, pero existen algunos (comunes entre los epifaunales, aunque también los hay infaunales) que lo hacen con una de las valvas contra el sustrato o de manera que la comisura queda aproximadamente paralela u oblicua a la interfase.Los pleurotéticos incluyen a todos los bivalvos cementantes, la mayoría de los apoyados, muchos bisados y algunos excavadores.Sin embargo estos hábitos no son ejercidos con exclusividad, presentándose frecuentes combinaciones de los mismos.Otras especies que viven fijas por el biso, pertenecientes a los géneros Isognomun y Barbatia, habitan introducidas a la manera de una cuña entre colonias de corales o rocas, igual que algunas especies nidificadoras libres.Muchas formas nadadoras, como los pectínidos, pasan buena parte de su existencia apoyadas en el fondo o fijas por el biso.
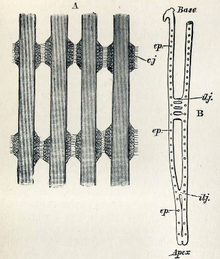
Cuatro filamentos de las branquias del mejillón azul (Mytilus edulis) a) parte de cuatro filamentos mostrando las uniones interfilamentosas ciliadas (cj) b) diagrama de un solo filamento mostrando las dos laminillas conectadas a intervalos por uniones interlamelares (ilj) y la posición de las uniones interfilamentosas ciliadas (cp).
Larva de bivalvo. Noreste del Mar Negro, aguas costeras a una profundidad de 0,5m.Larva de bivalvo. Noreste del Mar Negro, aguas costeras a una profundidad de 0,5m.