En fisiología visual , la adaptación es la capacidad de la retina del ojo para ajustarse a varios niveles de luz. La visión nocturna natural , o visión escotópica , es la capacidad de ver en condiciones de poca luz. En los humanos, las células bastón son exclusivamente responsables de la visión nocturna, ya que las células cono solo pueden funcionar en niveles de iluminación más altos. [1] La visión nocturna es de menor calidad que la visión diurna porque tiene una resolución limitada y no se pueden discernir los colores; solo se ven tonos de gris. [1] Para que los humanos pasen de la visión diurna a la nocturna, deben atravesar un período de adaptación a la oscuridad de hasta dos horas [2] en el que cada ojo se ajusta de una "configuración" de luminiscencia alta a una baja, lo que aumenta enormemente la sensibilidad, en muchos órdenes de magnitud. [1] Este período de adaptación es diferente entre las células bastón y cono y es el resultado de la regeneración de fotopigmentos para aumentar la sensibilidad de la retina. [1] La adaptación a la luz, en cambio, funciona muy rápidamente, en cuestión de segundos.
El ojo humano puede funcionar desde niveles de luz muy oscuros hasta niveles muy brillantes; sus capacidades de detección abarcan nueve órdenes de magnitud . Esto significa que la señal de luz más brillante y la más oscura que el ojo puede detectar están separadas por un factor de aproximadamente 1.000.000.000. Sin embargo, en un momento dado, el ojo solo puede detectar una relación de contraste de 1.000. Lo que permite un alcance más amplio es que el ojo adapta su definición de lo que es negro.
El ojo tarda aproximadamente entre 20 y 30 minutos en adaptarse por completo de la luz solar intensa a la oscuridad total y se vuelve entre 10 000 y 1 000 000 de veces más sensible que a plena luz del día. En este proceso, la percepción del color por parte del ojo también cambia (esto se denomina efecto Purkinje ). Sin embargo, el ojo tarda aproximadamente cinco minutos en adaptarse de la oscuridad a la luz solar intensa. Esto se debe a que los conos obtienen más sensibilidad cuando entran por primera vez en la oscuridad durante los primeros cinco minutos, pero los bastones toman el control después de cinco minutos o más. [3] Las células de los conos pueden recuperar la sensibilidad retiniana máxima en 9 o 10 minutos de oscuridad, mientras que los bastones necesitan entre 30 y 45 minutos para hacerlo. [4]
La adaptación a la oscuridad es mucho más rápida y profunda en los jóvenes que en los mayores. [5]
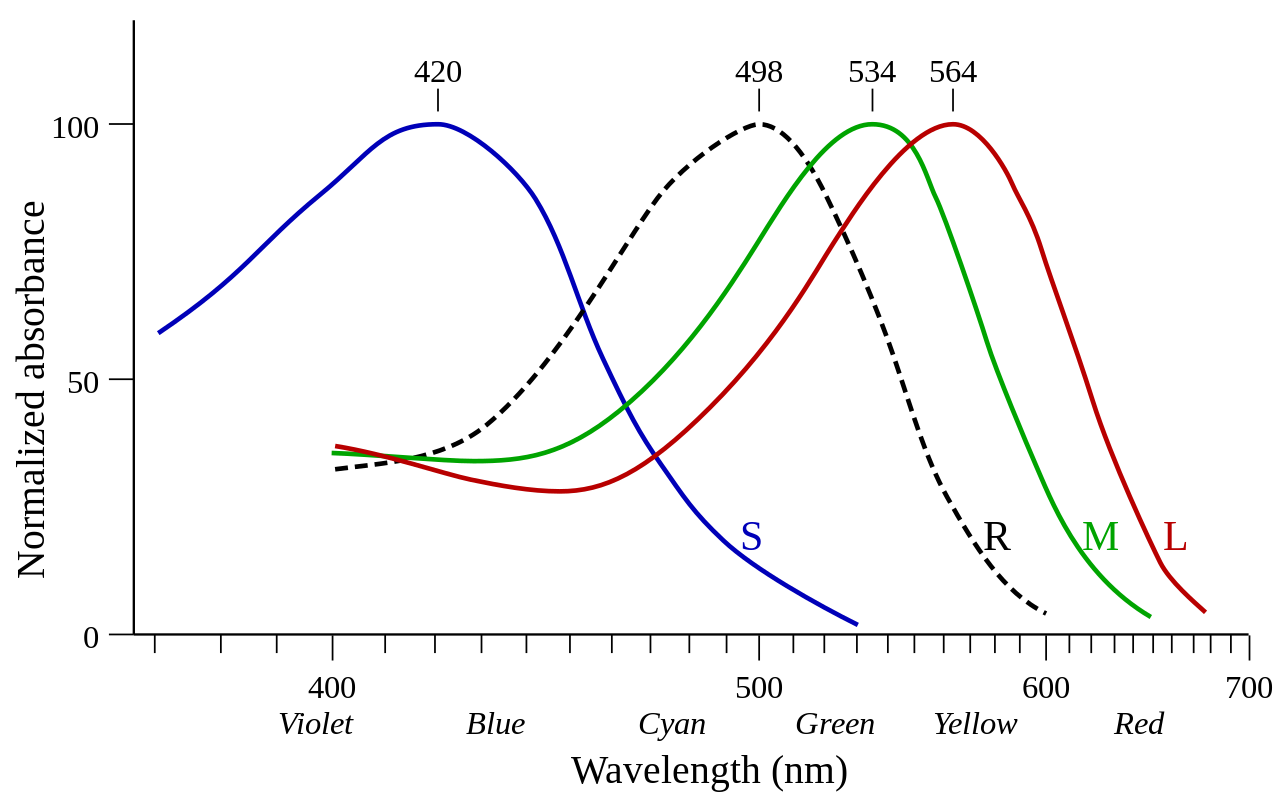
El ojo humano contiene tres tipos de fotorreceptores: bastones, conos y células ganglionares de la retina intrínsecamente fotosensibles (ipRGC). Los bastones y los conos son responsables de la visión y están conectados a la corteza visual. Las ipRGC están más conectadas con las funciones del reloj corporal y otras partes del cerebro, pero no con la corteza visual. Los bastones y los conos se pueden distinguir fácilmente por su estructura. Los fotorreceptores de cono tienen forma cónica y contienen opsinas de cono como pigmentos visuales. Existen tres tipos de fotorreceptores de cono, cada uno de los cuales es máximamente sensible a una longitud de onda específica de luz dependiendo de la estructura de su fotopigmento de opsina. [6] Las diversas células de cono son máximamente sensibles a longitudes de onda cortas (luz azul), longitudes de onda medias (luz verde) o longitudes de onda largas (luz roja). Los fotorreceptores de bastón solo contienen un tipo de fotopigmento, la rodopsina, que tiene una sensibilidad máxima a una longitud de onda de aproximadamente 500 nanómetros que corresponde a la luz azul-verde. [6] La distribución de las células fotorreceptoras a lo largo de la superficie de la retina tiene consecuencias importantes para la visión. [7] Los fotorreceptores de cono se concentran en una depresión en el centro de la retina conocida como fóvea central y disminuyen en número hacia la periferia de la retina. [7] Por el contrario, los fotorreceptores de bastón están presentes en alta densidad en la mayor parte de la retina con una marcada disminución en la fóvea. La percepción en entornos de alta luminiscencia está dominada por los conos a pesar del hecho de que son superados en gran medida por los bastones (aproximadamente 4,5 millones a 91 millones). [7]


Un mecanismo menor de adaptación es el reflejo pupilar a la luz , que ajusta muy rápidamente la cantidad de luz que llega a la retina en un factor de aproximadamente diez. Dado que contribuye solo a una pequeña fracción de la adaptación general a la luz, no se analiza más en este artículo.
En respuesta a los distintos niveles de luz ambiental, los bastones y conos del ojo funcionan tanto de forma aislada como en conjunto para ajustar el sistema visual. Los cambios en la sensibilidad de los bastones y conos del ojo son los principales factores que contribuyen a la adaptación a la oscuridad.
Por encima de un cierto nivel de luminancia (aproximadamente 0,03 cd/m2 ) , el mecanismo del cono interviene en la mediación de la visión; la visión fotópica . Por debajo de este nivel, el mecanismo de los bastones entra en juego proporcionando la visión escotópica (nocturna) . El rango en el que dos mecanismos trabajan juntos se denomina rango mesópico , ya que no hay una transición abrupta entre los dos mecanismos. Esta adaptación forma la base de la teoría de la duplicidad . [8]

Muchos animales, como los gatos, poseen una visión nocturna de alta resolución, lo que les permite discriminar objetos con frecuencias altas en entornos de baja iluminación. El tapetum lucidum es una estructura reflectante que es responsable de esta visión nocturna superior, ya que refleja la luz a través de la retina , exponiendo las células fotorreceptoras a una mayor cantidad de luz. [9] La mayoría de los animales que poseen un tapetum lucidum son nocturnos, probablemente porque al reflejar la luz a través de la retina, las imágenes iniciales se vuelven borrosas. [9] Los humanos, al igual que sus parientes primates, no poseen un tapetum lucidum y, por lo tanto, estaban predispuestos a ser una especie diurna. [10]
A pesar de que la resolución de la visión diurna humana es muy superior a la de la visión nocturna, la visión nocturna humana ofrece muchas ventajas. Al igual que muchos animales depredadores, los humanos pueden utilizar su visión nocturna para cazar y emboscar a otros animales sin darse cuenta. Además, en caso de que se produzca una situación de emergencia durante la noche, los humanos pueden aumentar sus posibilidades de supervivencia si son capaces de percibir su entorno y ponerse a salvo. Ambos beneficios pueden utilizarse para explicar por qué los humanos no perdieron por completo la capacidad de ver en la oscuridad de sus ancestros nocturnos. [11]

La rodopsina , un pigmento biológico en los fotorreceptores de la retina, se fotoblanquea inmediatamente en respuesta a la luz. [12] La fototransducción visual comienza con la isomerización del cromóforo del pigmento de 11-cis a todo-trans retinal . [13] Luego, este pigmento se disocia en opsina libre y todo-trans retinal. La adaptación a la oscuridad de los bastones y conos requiere la regeneración del pigmento visual a partir de opsina y 11-cis retinal. [13] Por lo tanto, el tiempo necesario para la adaptación a la oscuridad y la regeneración del pigmento está determinado en gran medida por la concentración local de 11-cis retinal y la velocidad a la que se entrega a la opsina en los bastones blanqueados. [14] La disminución en la entrada de iones de calcio después del cierre del canal provoca la fosforilación de la metarrodopsina II y acelera la inactivación de cis-retinal a trans-retinal. [13] La fosforilación de la rodopsina activada está mediada por la recoverina . [13] La regeneración de los fotopigmentos ocurre durante la adaptación a la oscuridad, aunque a velocidades marcadamente diferentes. [15] Los bastones son más sensibles a la luz y, por lo tanto, tardan más en adaptarse por completo al cambio de luz. Los bastones, cuyos fotopigmentos se regeneran más lentamente, no alcanzan su sensibilidad máxima hasta aproximadamente dos horas. [3] [16] Los conos tardan aproximadamente entre 9 y 10 minutos en adaptarse a la oscuridad. [3] La sensibilidad a la luz está modulada por cambios en los iones de calcio intracelulares y el monofosfato de guanosina cíclico . [17]
La sensibilidad de la vía de los bastones mejora considerablemente en un plazo de 5 a 10 minutos en la oscuridad. Se han utilizado pruebas de color para determinar el momento en el que el mecanismo de los bastones toma el control; cuando el mecanismo de los bastones toma el control, las manchas de color aparecen incoloras, ya que solo las vías de los conos codifican el color. [18]
Tres factores afectan la rapidez con la que el mecanismo de varilla se vuelve dominante:
En condiciones escotópicas , la concentración intracelular de cGMP es alta en los fotorreceptores. El cGMP se une a los canales de Na + regulados por cGMP y los abre para permitir la entrada de sodio y calcio. [22] La entrada de sodio contribuye a la despolarización, mientras que la entrada de calcio aumenta las concentraciones locales de calcio cerca del receptor. El calcio se une a una proteína moduladora, que se propone que sea GUCA1B , [23] eliminando el efecto estimulante de esta proteína sobre la guanilil ciclasa . [22] Esto reduce la producción de cGMP por la guanilil ciclasa para reducir la concentración de cGMP durante la oscuridad prolongada. La concentración elevada de calcio también aumenta la actividad de la fosfodiesterasa [22] que hidroliza el cGMP para reducir aún más su concentración. Esto reduce la apertura de los canales de Na + regulados por cGMP para hiperpolarizar la célula, volviéndola nuevamente sensible a pequeños aumentos en el brillo. Sin la adaptación a la oscuridad, el fotorreceptor permanecería despolarizado en condiciones escotópicas y, por lo tanto, tampoco respondería a pequeños cambios en el brillo.
La inhibición por parte de las neuronas también afecta a la activación en las sinapsis. Junto con el blanqueo del pigmento de los bastones o conos , se inhibe la fusión de señales en las células ganglionares , lo que reduce la convergencia.
La adaptación alfa, es decir , las fluctuaciones rápidas de sensibilidad, se alimentan del control nervioso. La fusión de señales en virtud de las células ganglionares difusas, así como de las células horizontales y amacrinas, permite un efecto acumulativo. Así, el área de estimulación es inversamente proporcional a la intensidad de la luz, un estímulo fuerte de 100 bastones equivale a un estímulo débil de 1.000 bastones.
En condiciones de luz suficientemente brillante, la convergencia es baja, pero durante la adaptación a la oscuridad, la convergencia de las señales de los bastones aumenta. Esto no se debe a cambios estructurales, sino a una posible interrupción de la inhibición que detiene la convergencia de los mensajes en condiciones de luz brillante. Si solo se abre un ojo, el ojo cerrado debe adaptarse por separado al volver a abrirse para coincidir con el ojo ya adaptado. [3]
Los oftalmólogos a veces miden la adaptación a la oscuridad de los pacientes utilizando un instrumento conocido como adaptómetro oscuro.
Existen varios adaptómetros oscuros comerciales (AdaptDx, MetroVision MonCvONE, Roland Consult DARK-Adaptometer). Además de estos dispositivos de visualización libre, recientemente se ha introducido un flujo de trabajo con seguimiento del fondo de ojo mediante la modificación de un dispositivo de microperimetría que permite realizar pruebas a pacientes con fijación inestable. [24]
Numerosos estudios clínicos han demostrado que la función de adaptación a la oscuridad se ve gravemente afectada desde las primeras etapas de la degeneración macular relacionada con la edad (DMRE), la retinosis pigmentaria (RP) y otras enfermedades de la retina, con un deterioro creciente a medida que progresan las enfermedades. [25] [26] La DMRE es una enfermedad crónica y progresiva que hace que una parte de la retina, llamada mácula, se deteriore lentamente con el tiempo. Es la principal causa de pérdida de visión entre las personas de 50 años o más. [27] Se caracteriza por una ruptura del complejo de membrana de Bruch/RPE en la retina, lo que lleva a una acumulación de depósitos de colesterol en la mácula. Con el tiempo, estos depósitos se convierten en drusas clínicamente visibles que afectan la salud de los fotorreceptores, causando inflamación y una predisposición a la neovascularización coroidea (NVC). Durante el curso de la enfermedad de DMRE, la función del EPR/Bruch continúa deteriorándose, lo que dificulta el transporte de nutrientes y oxígeno a los fotorreceptores de conos y bastones. Como efecto secundario de este proceso, los fotorreceptores muestran una adaptación a la oscuridad deteriorada porque necesitan estos nutrientes para la reposición de fotopigmentos y la eliminación de opsina para recuperar la sensibilidad escotópica después de la exposición a la luz.
La medición de la función de adaptación a la oscuridad de un paciente es, en esencia, un bioensayo de la salud de su membrana de Bruch. Por ello, las investigaciones han demostrado que, midiendo la adaptación a la oscuridad, los médicos pueden detectar la DMAE subclínica al menos tres años antes de que sea clínicamente evidente. [28]
.jpg/1280px-%D0%9F%D0%B5%D1%80%D1%81%D0%B5%D1%97%D0%B4%D0%B8-2021_-_%D0%B5%D0%BA%D1%81%D0%BA%D1%83%D1%80%D1%81%D1%96%D1%8F_%D0%93%D0%BE%D0%BB%D0%BE%D0%B2%D0%BD%D0%BE%D1%97_%D0%B0%D1%81%D1%82%D1%80%D0%BE%D0%BD%D0%BE%D0%BC%D1%96%D1%87%D0%BD%D0%BE%D1%97_%D0%BE%D0%B1%D1%81%D0%B5%D1%80%D0%B2%D0%B0%D1%82%D0%BE%D1%80%D1%96%D1%97_%D0%9D%D0%90%D0%9D_%D0%A3%D0%BA%D1%80%D0%B0%D1%97%D0%BD%D0%B8_(5).jpg)
Se ha demostrado que existen varios métodos diferentes, con distintos niveles de evidencia, que aumentan la velocidad a la que la visión puede adaptarse en la oscuridad.
Como las células bastón tienen una sensibilidad máxima en una longitud de onda de 500 nanómetros, no pueden percibir todos los colores del espectro visual. Debido a que las células bastón son insensibles a las longitudes de onda largas, el uso de luces rojas y gafas con lentes rojas se ha convertido en una práctica común para acelerar la adaptación a la oscuridad. [29] Para que la adaptación a la oscuridad se acelere significativamente, lo ideal es que una persona comience esta práctica 30 minutos antes de entrar en un entorno de baja luminiscencia. [30] Esta práctica permitirá que una persona mantenga su visión fotópica (diurna) mientras se prepara para la visión escotópica. La insensibilidad a la luz roja evitará que las células bastón se decoloren aún más y permitirá que el fotopigmento de rodopsina se recargue hasta su conformación activa. [29] Una vez que una persona entra en un entorno oscuro, la mayoría de sus células bastón ya se habrán adaptado a la oscuridad y podrán transmitir señales visuales al cerebro sin un período de acomodación. [30]
El concepto de lentes rojas para la adaptación a la oscuridad se basa en la experimentación de Antoine Béclère y sus primeros trabajos en radiología. En 1916, el científico Wilhelm Trendelenburg inventó el primer par de gafas de adaptación al rojo para que los radiólogos adaptaran sus ojos a la visualización en pantallas durante los procedimientos fluoroscópicos.
Aunque muchos aspectos del sistema visual humano siguen siendo inciertos, la teoría de la evolución de los fotopigmentos de bastones y conos es aceptada por la mayoría de los científicos. Se cree que los primeros pigmentos visuales fueron los de los fotorreceptores de conos, y que las proteínas opsinas de bastones evolucionaron más tarde. [31] Tras la evolución de los mamíferos a partir de sus ancestros reptiles hace aproximadamente 275 millones de años, hubo una fase nocturna en la que se perdió la visión compleja del color. [31] Como estos promamíferos eran nocturnos, aumentaron su sensibilidad en entornos de baja luminiscencia y redujeron su sistema fotópico de tetracromático a dicromático. [31] El cambio a un estilo de vida nocturno exigiría más fotorreceptores de bastones para absorber la luz azul emitida por la luna durante la noche. [32] Se puede extrapolar que la alta proporción de bastones a conos presente en los ojos humanos modernos se mantuvo incluso después del cambio de nocturno a diurno.
Se cree que la aparición de la tricromacia en los primates se produjo hace aproximadamente 55 millones de años, cuando la temperatura superficial del planeta empezó a aumentar. [31] Los primates eran de naturaleza diurna, no nocturna, y por tanto necesitaban un sistema visual fotópico más preciso. Era necesario un tercer fotopigmento cónico para cubrir todo el espectro visual, lo que permitía a los primates discriminar mejor entre las frutas y detectar las de mayor valor nutricional. [31]

La vitamina A es necesaria para el correcto funcionamiento del ojo humano. El fotopigmento rodopsina que se encuentra en los bastoncillos humanos está compuesto de retinal, una forma de vitamina A, unido a una proteína opsina. [35] Al absorber luz, la rodopsina se descompone en retinal y opsina mediante blanqueamiento. [35] La retina puede tener uno de dos destinos: puede recombinarse con la opsina para reformar la rodopsina o puede convertirse en retinol libre. [35] El científico estadounidense George Wald fue el primero en reconocer que el sistema visual gasta vitamina A y depende de la dieta para su reposición. [35]
La vitamina A cumple muchas funciones en el cuerpo humano, además de la visión saludable. Es vital para mantener un sistema inmunológico saludable y promover el crecimiento y el desarrollo normales. [36] El hombre y la mujer adultos promedio deben consumir 900 y 700 microgramos de vitamina A por día, respectivamente. [36] El consumo superior a 3000 microgramos por día puede provocar hipervitaminosis A aguda o crónica. [37]
La vitamina A está presente tanto en fuentes animales como vegetales en forma de retinoides y carotenoides, respectivamente. [36] Los retinoides pueden ser utilizados inmediatamente por el cuerpo tras su absorción en el sistema cardiovascular; sin embargo, los carotenoides de origen vegetal deben convertirse en retinol antes de que el cuerpo los utilice. [36] Las fuentes animales más ricas en vitamina A son el hígado, los productos lácteos y el pescado. [36] Las frutas y verduras que contienen grandes cantidades de carotenoides son de color verde oscuro, amarillo, naranja y rojo. [36]
Las proteínas opsinas basadas en vitamina A se han utilizado para detectar la luz en los organismos durante la mayor parte de la historia evolutiva, comenzando hace aproximadamente 3 mil millones de años. [38] Esta característica se ha transmitido de organismos unicelulares a organismos multicelulares, incluido el Homo sapiens. [38] Esta vitamina fue probablemente seleccionada por la evolución para detectar la luz porque la retina causa un cambio en la absorbancia de los fotorreceptores al rango de luz visible. [38] Este cambio en la absorbancia es especialmente importante para la vida en la Tierra porque generalmente coincide con la irradiancia máxima de la luz solar en su superficie. [38] Una segunda razón por la que la retina evolucionó para ser vital para la visión humana es porque sufre un gran cambio conformacional cuando se expone a la luz. [38] Se cree que este cambio conformacional hace que sea más fácil para la proteína fotorreceptora distinguir entre su estado silencioso y activado, controlando así mejor la fototransducción visual. [38]
Se han realizado varios estudios para probar la eficacia de la suplementación con vitamina A en la adaptación a la oscuridad. En un estudio de Cideciyan et al., se midió la duración de la adaptación a la oscuridad en un paciente con deficiencia sistémica de vitamina A (VAD) antes y después de la suplementación con vitamina A. [39] La función de adaptación a la oscuridad se midió antes de la suplementación, 1 día después del tratamiento y 75 días después del tratamiento. Se observó que después de solo un día de suplementación con vitamina A, la cinética de recuperación de la adaptación a la oscuridad se aceleró significativamente después del blanqueamiento de los fotorreceptores. [39] La adaptación a la oscuridad se aceleró aún más después de 75 días de tratamiento. [39]
Un estudio posterior de Kemp et al. estudió la adaptación a la oscuridad en sujetos con cirrosis biliar primaria y enfermedad de Crohn , ambos con deficiencia de vitamina A. [40] Dentro de los 8 días posteriores a la suplementación oral con vitamina A, ambos pacientes recuperaron su función visual a la normalidad. [40] Además, la cinética de adaptación mejoró significativamente en ambos sujetos después de la suplementación. [40]
Estudios independientes posteriores sobre la distrofia del fondo de ojo de Sorby confirmaron el efecto de la vitamina A en la adaptación a la oscuridad. [41] Asimismo, se demostró que la vitamina A acelera (en menor medida) la adaptación a la oscuridad en la DMAE. [42] [43]
Las antocianinas constituyen la mayoría de los 4000 fitoquímicos flavonoides conocidos. [44] Este grupo de aproximadamente 600 antioxidantes bioactivos tiene los efectos fisiológicos más fuertes de cualquier compuesto vegetal. [45] Estos químicos también son los más visibles de los fitoquímicos flavonoides porque proporcionan pigmentación azul, roja o púrpura brillante a muchas especies de plantas. [45] Las antocianinas también sirven para proteger los tejidos fotosintéticos de los rayos directos del sol. [46] Además, las propiedades antioxidantes , antiinflamatorias y vasoprotectoras de las antocianinas les permiten demostrar diversos efectos sobre la salud. [45] En los humanos, las antocianinas son efectivas para una variedad de condiciones de salud que incluyen daño neurológico, aterosclerosis, diabetes y discapacidad visual. [46] Las antocianinas interactúan frecuentemente con otros fitoquímicos para potenciar los efectos biológicos; por lo tanto, las contribuciones de las biomoléculas individuales siguen siendo difíciles de descifrar. [44] Como resultado de que las antocianinas proporcionan una coloración brillante a las flores, las plantas que contienen estos fitoquímicos tienen un éxito natural en la atracción de polinizadores como pájaros y abejas. [46] Las frutas y verduras producidas por estas plantas también tienen una pigmentación brillante que atrae a los animales para que las coman y dispersen las semillas. [46] Debido a este mecanismo natural, las plantas que contienen antocianinas son muy abundantes en la mayoría de las áreas del mundo. La alta abundancia y distribución de plantas que contienen antocianinas las convierten en una fuente natural de alimento para muchos animales. A través de evidencia fósil se sabe que estos compuestos fueron consumidos en grandes cantidades por los homínidos primitivos. [45]
Durante la Primera y la Segunda Guerra Mundial, los aviadores de la Fuerza Aérea Británica consumían grandes cantidades de mermelada de arándanos. Los aviadores consumían este alimento rico en antocianinas debido a sus numerosos beneficios visuales, incluida la adaptación acelerada a la oscuridad, lo que sería valioso para las misiones de bombardeo nocturno. [47]

Las frutas y verduras de colores vivos son ricas en antocianinas. Esto tiene sentido intuitivamente porque las antocianinas aportan pigmentación a las plantas. Las moras son los alimentos más ricos en antocianinas, ya que contienen entre 89 y 211 miligramos por cada 100 gramos. [46] Otros alimentos ricos en este fitoquímico son las cebollas rojas, los arándanos azules, los arándanos rojos, la col lombarda y la berenjena. [46] La ingestión de cualquiera de estas fuentes de alimentos producirá una variedad de fitoquímicos además de antocianinas porque existen juntas de forma natural. [44] Se estima que la ingesta diaria de antocianinas es de aproximadamente 200 miligramos en un adulto medio; sin embargo, este valor puede alcanzar varios gramos al día si una persona consume suplementos de flavonoides. [44]
Las antocianinas aceleran la adaptación a la oscuridad en los seres humanos al mejorar la regeneración del fotopigmento de los bastones, la rodopsina. [48] Las antocianinas logran esto al unirse directamente a la opsina tras la degradación de la rodopsina a sus componentes individuales por la luz. [48] Una vez unida a la opsina, la antocianina cambia su estructura, acelerando así su acceso al bolsillo de unión de la retina. Al tener una dieta rica en antocianinas, un individuo puede generar rodopsina en períodos de tiempo más cortos debido a la mayor afinidad de la opsina con la retina. [48] A través de este mecanismo, un individuo puede acelerar la adaptación a la oscuridad y lograr la visión nocturna en un período de tiempo más corto.
En un estudio doble ciego controlado con placebo realizado por Nakaishi et al., se proporcionó a varios participantes un concentrado de antocianina en polvo derivado de grosellas negras. [49] [ ¿fuente médica poco confiable? ] Los participantes recibieron una de tres dosis de antocianinas para medir si el resultado se produjo de manera dependiente de la dosis. El período de adaptación a la oscuridad se midió antes y dos horas después de la suplementación en todos los participantes. Los resultados de este experimento indican que las antocianinas aceleraron significativamente la adaptación a la oscuridad con solo un nivel de dosis en comparación con el placebo. [49] [ ¿fuente médica poco confiable? ] Al observar los datos en su conjunto, Nakaishi et al. concluyeron que las antocianinas redujeron efectivamente el período de adaptación a la oscuridad de manera dependiente de la dosis. [49] [ ¿fuente médica poco confiable? ]
A pesar de que muchos científicos creen que las antocianinas son beneficiosas para acelerar la adaptación a la oscuridad en los seres humanos, un estudio realizado por Kalt et al. en 2014 demostró que las antocianinas de los arándanos no tienen ningún efecto. En este estudio, se realizaron dos estudios doble ciego controlados con placebo para examinar la adaptación a la oscuridad después de la ingesta de productos de arándanos. [50] En ninguno de los estudios, la ingesta de antocianinas de arándanos afectó la duración de la adaptación a la oscuridad. [50] A partir de estos resultados, Kalt et al. concluyeron que las antocianinas de los arándanos no aportan ninguna diferencia significativa al componente de adaptación a la oscuridad de la visión humana. [50]
En la adaptación a la luz, el ojo debe adaptarse rápidamente a la iluminación del fondo para poder distinguir los objetos que se encuentran en él. El proceso de adaptación a la luz se lleva a cabo en un período de cinco minutos.
La reacción fotoquímica es:

Mediante experimentos de umbral de incremento, la adaptación a la luz se puede medir clínicamente. [51] En un experimento de umbral de incremento, se presenta un estímulo de prueba sobre un fondo de una cierta luminancia , el estímulo se aumenta hasta que se alcanza el umbral de detección contra el fondo. A través de este método se obtiene una curva de umbral versus intensidad TVI monofásica o bifásica tanto para conos como para bastones .
Cuando se toma de forma aislada la curva de umbral de un único sistema (es decir, sólo conos o sólo bastones ), se puede observar que posee cuatro secciones: [52]

La insuficiencia de adaptación se presenta más comúnmente como una adaptación insuficiente a un entorno oscuro, llamada ceguera nocturna o nictalopía . [35] El problema opuesto, conocido como hemeralopía , es decir, la incapacidad de ver claramente en luz brillante, es mucho más raro.
La fóvea es ciega a la luz tenue (debido a su disposición de solo conos) y los bastones son más sensibles, por lo que una estrella tenue en una noche sin luna debe verse de lado , de modo que estimule los bastones. Esto no se debe al ancho de la pupila, ya que una pupila artificial de ancho fijo da los mismos resultados. [3]
La ceguera nocturna puede ser causada por varios factores, el más común de los cuales es la deficiencia de vitamina A. Si se detecta a tiempo, la nictalopía se puede revertir y se puede recuperar la función visual; sin embargo, la deficiencia prolongada de vitamina A puede provocar una pérdida visual permanente si no se trata. [55]
La ceguera nocturna es especialmente frecuente en los países en desarrollo debido a la desnutrición y, por lo tanto, a la falta de vitamina A en la dieta. [55] En los países desarrollados, la ceguera nocturna ha sido históricamente poco común debido a la disponibilidad adecuada de alimentos; sin embargo, se espera que la incidencia aumente a medida que la obesidad se vuelva más común. El aumento de las tasas de obesidad corresponde a un mayor número de cirugías bariátricas, lo que provoca una mala absorción de vitamina A en el cuerpo humano. [55]