La memoria de reconocimiento , una subcategoría de la memoria explícita , es la capacidad de reconocer eventos, objetos o personas encontrados previamente. [1] Cuando se vuelve a experimentar el evento experimentado anteriormente, este contenido ambiental se compara con representaciones de memoria almacenadas, generando señales coincidentes. [2] Como lo establecieron por primera vez experimentos de psicología en la década de 1970, la memoria de reconocimiento de imágenes es bastante notable: los humanos pueden recordar miles de imágenes con gran precisión después de ver cada una de ellas solo una vez y solo durante unos segundos. [3]
La memoria de reconocimiento se puede subdividir en dos procesos componentes: recuerdo y familiaridad, a veces denominados "recordar" y "saber", respectivamente. [1] El recuerdo es la recuperación de detalles asociados con el evento experimentado previamente. Por el contrario, la familiaridad es la sensación de que el evento fue experimentado previamente, sin recuerdo. Por tanto, la distinción fundamental entre los dos procesos es que el recuerdo es un proceso de búsqueda lento y controlado, mientras que la familiaridad es un proceso rápido y automático. [4] [5]
Ejemplo del "carnicero en el autobús" de Mandler: [4]
Imagínese sentarse en un autobús lleno de gente. Miras a tu izquierda y notas a un hombre. Inmediatamente, te invade la sensación de que has visto a este hombre antes, pero no puedes recordar quién es. Este sentimiento provocado automáticamente es el de familiaridad. Mientras intentas recordar quién es este hombre, comienzas a recuperar detalles específicos sobre tu encuentro anterior. Por ejemplo, quizás recuerdes que este hombre te entregó una fina chuleta de carne en el supermercado. O quizás lo recuerdes usando un delantal. Este proceso de búsqueda es el recuerdo.
El fenómeno de la familiaridad y el reconocimiento se describe desde hace mucho tiempo en libros y poemas. Dentro del campo de la Psicología , la memoria de reconocimiento fue aludida por primera vez por Wilhelm Wundt en su concepto de conocimiento de nuevo o asimilación de una imagen de memoria anterior a una nueva. El primer intento formal de describir el reconocimiento fue realizado por el doctor inglés Arthur Wigan en su libro Duality of the Mind . Aquí describe los sentimientos de familiaridad que experimentamos debido a que el cerebro es un órgano doble . [6] En esencia, percibimos cosas con la mitad de nuestro cerebro y si de alguna manera se pierden en la traducción al otro lado del cerebro, esto causa la sensación de reconocimiento cuando volvemos a ver dicho objeto, persona, etc. Sin embargo, asumió incorrectamente que estos sentimientos ocurren sólo cuando la mente está agotada (por hambre, falta de sueño, etc.). Su descripción, aunque elemental en comparación con los conocimientos actuales, sentó las bases y despertó el interés por este tema en investigadores posteriores. Arthur Allin (1896) fue la primera persona en publicar un artículo que intentaba definir y diferenciar explícitamente entre definiciones subjetivas y objetivas de la experiencia de reconocimiento, aunque sus hallazgos se basan principalmente en introspecciones . Allin corrige la noción de Wigan de mente agotada afirmando que este estado de semi-sueño no es el proceso de reconocimiento. [6] Se refiere bastante brevemente a los correlatos fisiológicos de este mecanismo como si tuvieran que ver con la corteza, pero no entra en detalles sobre dónde se encuentran estos sustratos. [6] Su explicación objetiva de la falta de reconocimiento es cuando una persona observa un objeto por segunda vez y experimenta la sensación de familiaridad que experimentó con este objeto en un momento anterior. [6] Woodsworth (1913) y Margaret y Edward Strong (1916) fueron las primeras personas en utilizar y registrar experimentalmente los hallazgos empleando la tarea de coincidencia retardada con la muestra para analizar la memoria de reconocimiento. [7] A continuación, Benton Underwood fue la primera persona en analizar el concepto de errores de reconocimiento en relación con las palabras en 1969. Descifró que estos errores de reconocimiento ocurren cuando las palabras tienen atributos similares. [8] Luego vinieron los intentos de determinar los límites superiores de la memoria de reconocimiento, una tarea que emprendió Standing (1973). Determinó que la capacidad de fotografías es casi ilimitada. [9] En 1980, George Mandler introdujo la distinción recuerdo-familiaridad, más formalmente conocida como teoría del proceso dual [4]
Es discutible si la familiaridad y el recuerdo deben considerarse categorías separadas de la memoria de reconocimiento. Esta distinción entre familiaridad y recuerdo es lo que se denomina modelo/teoría de proceso dual . "A pesar de la popularidad y la influencia de las teorías de proceso dual [para la memoria de reconocimiento], son controvertidas debido a la dificultad de obtener estimaciones empíricas separadas de recuerdo y familiaridad y la mayor parsimonia asociada con las teorías de proceso único". [10] Una crítica común a los modelos de reconocimiento de proceso dual es que el recuerdo es simplemente una versión más fuerte (es decir, más detallada o vívida) de la familiaridad. Así, en lugar de consistir en dos categorías separadas, los modelos de proceso único consideran la memoria de reconocimiento como un continuo que va desde los recuerdos débiles hasta los recuerdos fuertes. [1] Un relato de la historia de los modelos de proceso dual desde finales de la década de 1960 también incluye técnicas para la medición de los dos procesos. [11]
La evidencia de la visión de proceso único proviene de un estudio de registro de electrodos realizado en pacientes epilépticos que realizaron una tarea de reconocimiento de elementos. [12] Este estudio encontró que las neuronas del hipocampo, independientemente del recuerdo exitoso, respondían a la familiaridad de los objetos. Por lo tanto, es posible que el hipocampo no sirva exclusivamente para el proceso de recuerdo. Sin embargo, también descubrieron que el reconocimiento exitoso de elementos no estaba relacionado con si las neuronas de "familiaridad" se activaban o no. Por lo tanto, no está del todo claro qué respuestas se relacionan con el reconocimiento exitoso del ítem. Sin embargo, un estudio sugirió que la activación del hipocampo no significa necesariamente que se producirá un recuerdo consciente. [13] En este estudio de reconocimiento asociativo de escena de objeto, la activación del hipocampo no se relacionó con el recuerdo asociativo exitoso; Sólo cuando se activaron la corteza prefrontal y el hipocampo se observó un desempeño exitoso. Además, la evidencia del seguimiento ocular reveló que los participantes miraban durante más tiempo el estímulo correcto, y esto se relacionaba con aumentos en la actividad del hipocampo. Por lo tanto, el hipocampo puede desempeñar un papel en la recuperación de información relacional, pero requiere activación concomitante con la corteza prefrontal para el recuerdo consciente.
Los estudios con amnésicos no parecen apoyar la noción de proceso único. Varios informes presentan a pacientes con daño selectivo en el hipocampo que sólo tienen problemas de recuerdo pero no de familiaridad, lo que proporciona un apoyo tentativo a los modelos de proceso dual. [14] Además, se ha observado una doble disociación entre recuerdo y familiaridad. [15] A la paciente NB se le extirparon regiones de sus lóbulos temporales mediales, incluidas la corteza perirrinal y la corteza entorrinal, pero se salvaron el hipocampo y la corteza parahipocámpica. Mostró familiaridad deteriorada pero procesos de recuerdo intactos en relación con los controles en un paradigma de reconocimiento de sí-no, y esto se aclaró mediante procedimientos ROC, RK y fecha límite de respuesta. En otro estudio, incluso cuando el rendimiento entre los pacientes NB se comparó con el de un paciente amnésico al que se le extirpó el hipocampo, la doble disociación todavía estaba presente. [16] Si bien el rendimiento se igualó post hoc y se necesita replicación, esta evidencia descarta la idea de que estas regiones del cerebro sean parte de un sistema unitario de fortaleza de la memoria. [17] En cambio, esta doble disociación sugiere fuertemente que distintas regiones y sistemas cerebrales subyacen a los procesos de recuerdo y familiaridad.
Las teorías del proceso dual permiten distinguir dos tipos de reconocimiento: primero, reconocer QUE uno ha encontrado algún objeto/evento antes; y segundo reconocer QUÉ fue ese objeto/evento. Así, uno puede reconocer un rostro, pero sólo después recordar de quién era. [11] El reconocimiento retardado también muestra diferencias entre los procesos de familiaridad rápida y de recuerdo lento [18] [19] Además, en el sistema de “familiaridad” de reconocimiento de la memoria se distinguen dos subsistemas funcionales: el primero es responsable del reconocimiento de estímulos presentados previamente. y el segundo admite el reconocimiento de objetos como nuevos. [20]
Actualmente, la investigación neurocientífica no ha proporcionado una respuesta definitiva a esta controversia, aunque favorece en gran medida los modelos de proceso dual. Si bien muchos estudios proporcionan evidencia de que el recuerdo y la familiaridad están representados en regiones separadas del cerebro, otros estudios muestran que no siempre es así; Puede haber una gran superposición neuroanatómica entre los dos procesos. [1] A pesar de que la familiaridad y el recuerdo a veces activan las mismas regiones del cerebro, normalmente son bastante distintas funcionalmente. [1]
La cuestión de si el recuerdo y la familiaridad existen como dos categorías independientes o a lo largo de un continuo puede, en última instancia, ser irrelevante; La conclusión es que la distinción entre recuerdo y familiaridad ha sido extremadamente útil para comprender cómo funciona la memoria de reconocimiento.
Se utiliza para evaluar la memoria de reconocimiento según el patrón de respuestas de sí o no. [21] Esta es una de las formas más simples de prueba de reconocimiento y se realiza dándole a un participante un elemento y pidiéndole que indique "sí" si es antiguo o "no" si es un elemento nuevo. Este método de prueba de reconocimiento hace que el proceso de recuperación sea fácil de registrar y analizar. [22]
Se pide a los participantes que identifiquen cuál de varios ítems (dos a cuatro) es correcto. [21] Uno de los elementos presentados es el objetivo: un elemento presentado previamente. Los demás elementos son similares y actúan como distractores. Esto permite al experimentador cierto grado de manipulación y control en la similitud o semejanza de los elementos. Esto ayuda a proporcionar una mejor comprensión de la recuperación y qué tipos de conocimiento existente utilizan las personas para decidir basándose en la memoria. [21]
Cuando se registra el tiempo de respuesta (en milisegundos o segundos), se cree que una velocidad más rápida refleja un proceso más simple, mientras que los tiempos más lentos reflejan procesos fisiológicos más complejos. [21]
Hermann von Helmholtz fue el primer psicólogo que se preguntó si la velocidad de un impulso nervioso podía ser mensurable. [23] Ideó un sistema experimental para medir procesos psicológicos con una escala de tiempo muy precisa y crítica. El nacimiento de la cronometría mental se atribuye a un experimento del colega de Helmholtz, Franciscus Donders . En el experimento, colocó electrodos en ambos pies del sujeto. Luego administró una leve descarga al pie izquierdo o derecho y le dijo al sujeto que moviera la mano del mismo lado, lo que desactivó el estímulo (la descarga). En una condición diferente, al sujeto no se le dijo en qué pie actuaría el estímulo. La diferencia de tiempo entre estas condiciones se midió como un quinceavo de segundo. Este fue un hallazgo significativo en la psicología experimental temprana, porque los investigadores anteriormente pensaban que los procesos psicológicos eran demasiado rápidos para medirlos. [23]
El modelo de Atkinson y Juola (1973) sugirió un modelo temprano de teorías de procesos duales. [24] En esta teoría, el proceso de familiaridad sería el primero activado como una búsqueda rápida de reconocimiento. Si esto no logra recuperar el rastro de la memoria, entonces hay una búsqueda más forzada en el almacén de la memoria a largo plazo. [24]
El modelo de la "carrera de caballos" es una visión más reciente de las teorías del proceso dual. Este punto de vista sugiere que los dos procesos de familiaridad y recuerdo ocurren simultáneamente, pero que la familiaridad, al ser el proceso más rápido, completa la búsqueda antes del recuerdo. [4] Este punto de vista es cierto: la idea de que la familiaridad es un proceso inconsciente, mientras que el recuerdo es más consciente y reflexivo.
En circunstancias de incertidumbre, la identificación de un suceso anterior depende de los procesos de toma de decisiones . La información disponible debe compararse con algunos criterios internos que orienten sobre qué decisión es más ventajosa. [25]
La teoría de la detección de señales se ha aplicado a la memoria de reconocimiento como método para estimar el efecto de la aplicación de estos criterios internos, denominado sesgo . Es fundamental para el modelo de proceso dual la suposición de que la memoria de reconocimiento refleja un proceso de detección de señales en el que los elementos antiguos y nuevos tienen cada uno una distribución distinta a lo largo de una dimensión, como la familiaridad. [26] La aplicación de la Teoría de Detección de Señales (SDT) a la memoria depende de concebir un rastro de memoria como una señal que el sujeto debe detectar para poder realizar una tarea de retención. Dada esta concepción del rendimiento de la memoria, es razonable suponer que los porcentajes de puntuaciones correctas pueden ser indicadores sesgados de retención (al igual que los umbrales pueden ser indicadores sesgados del rendimiento sensorial) y, además, que se deben utilizar técnicas de SDT siempre que sea posible para separar los resultados. Aspectos verdaderamente basados en la retención del rendimiento de la memoria desde los aspectos de decisión. [27] En particular, asumimos que el sujeto compara la fuerza traza del ítem de prueba con un criterio, respondiendo "sí" si la fuerza excede el criterio y "no" en caso contrario. Hay dos tipos de ítems de prueba, "antiguo" (un ítem de prueba que apareció en la lista de esa prueba) y "nuevo" (uno que no apareció en la lista). La teoría de la fuerza supone que puede haber ruido en el valor de la fuerza de la traza, la ubicación del criterio, o ambos. Suponemos que este ruido se distribuye normalmente. [28] El criterio de informe puede cambiar a lo largo del continuo en la dirección de más aciertos falsos, o más fallos. La fuerza de la memoria momentánea de un ítem de prueba se compara con los criterios de decisión y si la fuerza del ítem cae dentro de la categoría de juicio, Jt, definida por la ubicación de los criterios, emite el juicio S. Se supone que la fuerza de un ítem disminuye monótonamente (con cierto error). varianza) como una función continua del tiempo o del número de elementos intermedios. [29] Los aciertos falsos son palabras "nuevas" incorrectamente reconocidas como antiguas, y una mayor proporción de ellas representa un sesgo liberal. [25] Los errores son palabras "viejas" erróneamente no se reconocen como antiguos y una mayor proporción de ellos representa un sesgo conservador. [25] Las distribuciones relativas de aciertos y errores falsos se pueden utilizar para interpretar el desempeño de la tarea de reconocimiento y corregir las conjeturas. [30] Solo los elementos objetivo pueden generar una respuesta de reconocimiento por encima del umbral porque solo ellos aparecieron en la lista. Los señuelos, junto con los objetivos olvidados, caen por debajo del umbral, lo que significa que no generan ninguna señal de memoria. Las falsas alarmas en este modelo reflejan conjeturas sin memoria que se hacen sobre algunos de los señuelos. [31]
El nivel de procesamiento cognitivo realizado ante un estímulo determinado tiene un efecto en el rendimiento de la memoria de reconocimiento, y un procesamiento asociativo más elaborado da como resultado un mejor rendimiento de la memoria. [32] Por ejemplo, el rendimiento del reconocimiento se mejora mediante el uso de asociaciones semánticas en lugar de asociaciones de características . [33] Sin embargo, este proceso está mediado por otras características de los estímulos, por ejemplo, la relación de los elementos entre sí. Si los elementos están altamente interrelacionados, el procesamiento específico de elementos de menor profundidad (como calificar lo agradable de cada elemento) ayuda a distinguirlos entre sí y mejora el rendimiento de la memoria de reconocimiento sobre el procesamiento relacional. [34] Este fenómeno inusual se explica por la tendencia automática a realizar procesamiento relacional en elementos altamente interrelacionados. El rendimiento del reconocimiento mejora mediante un procesamiento adicional, incluso de un nivel más bajo de asociatividad, pero no mediante una tarea que duplique el procesamiento automatizado ya realizado en la lista de elementos. [35]
Hay una variedad de formas en que el contexto puede influir en la memoria. La especificidad de codificación describe cómo se mejora el rendimiento de la memoria si las condiciones de prueba coinciden con las condiciones de aprendizaje (codificación). [36] Ciertos aspectos durante el período de aprendizaje, ya sea el entorno, su estado físico actual o incluso su estado de ánimo, quedan codificados en el rastro de la memoria. Más adelante, durante la recuperación, cualquiera de estos aspectos puede servir como clave para ayudar en el reconocimiento. Por ejemplo, la investigación de Godden y Baddeley [37] probó este concepto en buceadores. Un grupo de buzos aprendió listas de palabras en tierra y otro grupo aprendió bajo el agua. No es sorprendente que los resultados de las pruebas fueran más altos cuando las condiciones de recuperación coincidían con las condiciones de codificación (aquellos que aprendieron las listas de palabras en tierra obtuvieron mejores resultados en tierra, y aquellos que aprendieron bajo el agua obtuvieron mejores resultados bajo el agua). También se han realizado estudios que muestran efectos similares con respecto al estado físico de un individuo. Esto se conoce como aprendizaje dependiente del estado . [38] Otro tipo de especificidad de codificación es la memoria congruente con el estado de ánimo, donde es más probable que los individuos recuerden material si el contenido emocional del material y el estado de ánimo predominante en el momento del recuerdo coinciden. [39]
La presencia de otros individuos también puede influir en el reconocimiento. Dos efectos opuestos, la inhibición colaborativa y la facilitación colaborativa, impactan el rendimiento de la memoria en grupos. Específicamente, la facilitación colaborativa se refiere al mayor desempeño en tareas de reconocimiento en grupos. Lo contrario, la inhibición colaborativa, se refiere a una disminución del rendimiento de la memoria en tareas de recuerdo en grupos. [40] Esto se debe a que en una tarea de recuerdo , se debe activar un rastro de memoria específico, y las ideas externas podrían producir una especie de interferencia. El reconocimiento, por otro lado, no utiliza el mismo plan de recuperación que el recuerdo y, por lo tanto, no se ve afectado. [41]
Las dos categorías básicas de errores de memoria de reconocimiento son aciertos falsos (o falsas alarmas) y errores. [30] Un resultado falso es la identificación de un suceso como antiguo cuando en realidad es nuevo. Un error es no identificar un suceso anterior como antiguo.
Dos tipos específicos de aciertos falsos surgen cuando se obtienen mediante el uso de un señuelo de reconocimiento. El primero es un error de característica, en el que una parte de un estímulo antiguo se presenta en combinación con un elemento nuevo. [42] Por ejemplo, si la lista original contenía "mirlo, cebo de cárcel, trigo sarraceno", se puede provocar un error de característica mediante la presentación de "perdigones" o "chantaje" en la prueba, ya que cada uno de estos señuelos tiene un viejo y un nuevo componente. [43] El segundo tipo de error es un error de conjunción, en el que se combinan partes de múltiples estímulos antiguos. [42] Usando el mismo ejemplo, "jailbird" podría provocar un error de conjunción, ya que es una conjunción de dos estímulos antiguos. [43] Ambos tipos de errores pueden provocarse a través de modalidades auditivas y visuales , lo que sugiere que los procesos que producen estos errores no son específicos de la modalidad. [44]
Se puede inducir un tercer error de acierto falso mediante el uso del paradigma de Deese-Roediger-McDermott [45] . Si todos los elementos estudiados están muy relacionados con una palabra que no aparece en la lista, es muy probable que el sujeto reconozca esa palabra como antigua en la prueba. [46] Un ejemplo de esto sería una lista que contenga las siguientes palabras: siesta, somnoliento, cama, edredón, noche, relax. El atractivo en este caso es la palabra "dormir". Es muy probable que se reconozca falsamente que 'dormir' aparece en esa lista debido al nivel de activación recibido de las palabras de la lista. Este fenómeno es tan generalizado que la tasa de respuestas falsas generadas de esta manera puede incluso superar la tasa de respuestas correctas [47]
Según Robert L. Green (1996), el efecto espejo se produce cuando estímulos que son fáciles de reconocer como viejos cuando son viejos también lo son fácilmente como nuevos cuando son nuevos en el reconocimiento. El efecto espejo se refiere a la consistencia del reconocimiento de los estímulos en la memoria. [ se necesita aclaración ] En otras palabras, son más fáciles de recordar cuando has estudiado previamente los estímulos, es decir, antiguos, y más fáciles de rechazar cuando no los has visto antes, es decir, nuevos. Murray Glanzer y John K. Adams describieron por primera vez el efecto espejo en 1985. El efecto espejo ha sido eficaz en pruebas de reconocimiento asociativo, medidas de respuestas de latencia, discriminaciones de orden y otras (Glanzer & Adams, 1985).
En general, la investigación sobre los sustratos neuronales de la familiaridad y el recuerdo demuestra que estos procesos típicamente involucran diferentes regiones del cerebro, apoyando así una teoría de proceso dual de la memoria de reconocimiento. Sin embargo, debido a la complejidad y la interconectividad inherente de las redes neuronales del cerebro, y dada la estrecha proximidad de las regiones involucradas en la familiaridad con las regiones involucradas en el recuerdo, es difícil identificar las estructuras que están específicamente relacionadas con el recuerdo o con la familiaridad. Lo que se sabe actualmente es que la mayoría de las regiones neuroanatómicas implicadas en la memoria de reconocimiento están asociadas principalmente con un subcomponente más que con el otro.
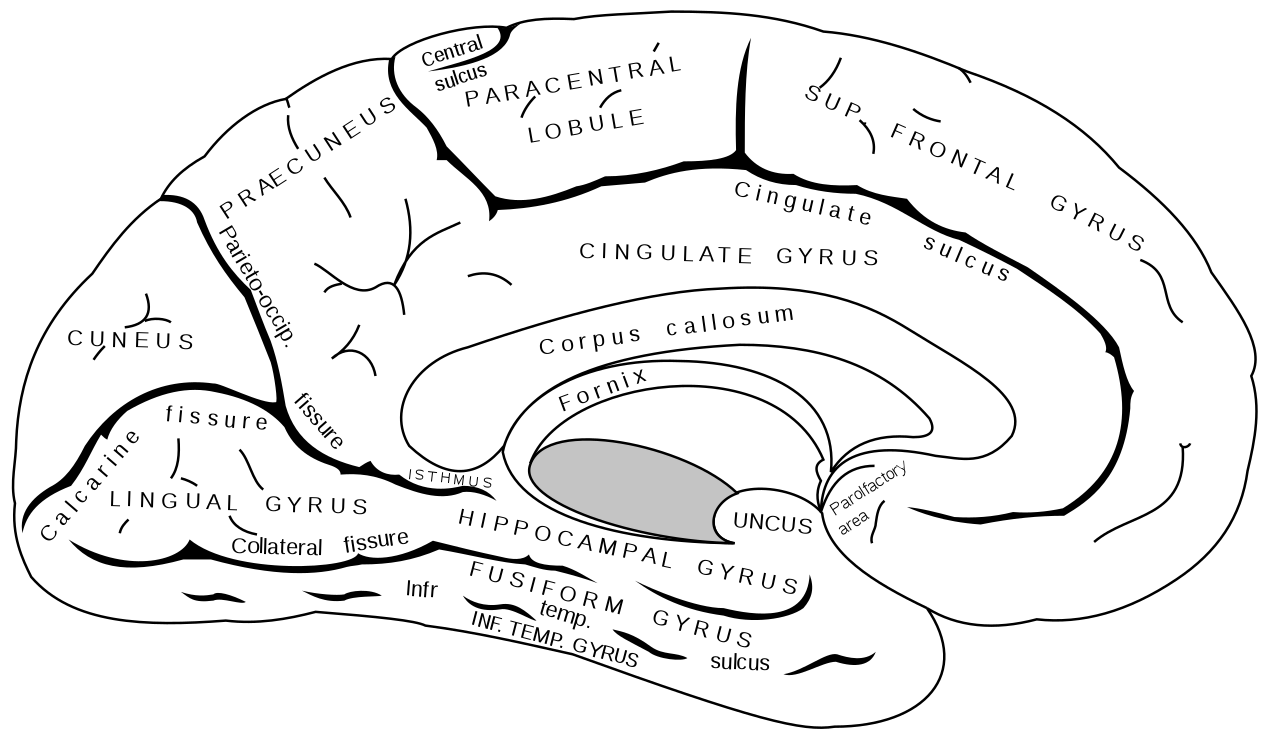
La memoria de reconocimiento depende de manera crítica de una red organizada jerárquicamente de áreas cerebrales que incluyen la corriente ventral visual , las estructuras del lóbulo temporal medial , el lóbulo frontal y las cortezas parietales [48] junto con el hipocampo . [49] Como se mencionó anteriormente, los procesos de recuerdo y familiaridad se representan de manera diferente en el cerebro. Como tal, cada una de las regiones enumeradas anteriormente se puede subdividir según qué parte esté involucrada principalmente en el recuerdo o en la familiaridad. En la corteza temporal, por ejemplo, la región medial está relacionada con el recuerdo, mientras que la región anterior está relacionada con la familiaridad. De manera similar, en la corteza parietal, la región lateral está relacionada con el recuerdo, mientras que la región superior está relacionada con la familiaridad. [49] Una explicación aún más específica divide la región parietal medial, relacionando el cingulado posterior con el recuerdo y el precuneus con la familiaridad. [49] El hipocampo desempeña un papel destacado en el recuerdo, mientras que la familiaridad depende en gran medida de las regiones medial-temporales circundantes, especialmente la corteza perirrinal. [50] Finalmente, aún no está claro qué regiones específicas de los lóbulos prefrontales están asociadas con el recuerdo versus la familiaridad, aunque hay evidencia de que la corteza prefrontal izquierda se correlaciona más fuertemente con el recuerdo, mientras que la corteza prefrontal derecha está más involucrada en la familiaridad. [51] [52] Aunque originalmente se planteó la hipótesis de que la activación del lado izquierdo involucrada en el recuerdo era el resultado del procesamiento semántico de las palabras (muchos de estos estudios anteriores utilizaron palabras escritas para los estímulos), estudios posteriores que utilizaron estímulos no verbales produjeron el mismo hallazgo, lo que sugiere que la activación prefrontal en el hemisferio izquierdo resulta de cualquier tipo de recuerdo detallado. [53]
Como se mencionó anteriormente, la memoria de reconocimiento no es un concepto independiente; más bien es un subsistema de memoria altamente interconectado e integrado. Quizás de manera engañosa, las regiones del cerebro enumeradas anteriormente corresponden a una comprensión abstracta y altamente generalizada de la memoria de reconocimiento, en la que los estímulos o elementos a reconocer no están especificados. En realidad, sin embargo, la localización de la activación cerebral implicada en el reconocimiento depende en gran medida de la naturaleza del estímulo mismo. Considere las diferencias conceptuales entre el reconocimiento de palabras escritas y el reconocimiento de rostros humanos. Se trata de dos tareas cualitativamente diferentes y, como tales, no sorprende que impliquen regiones adicionales y distintas del cerebro. Reconocer palabras, por ejemplo, implica el área de formación de palabras visuales , una región en la circunvolución fusiforme izquierda, que se cree que se especializa en reconocer palabras escritas. [54] Del mismo modo, la zona fusiforme del rostro , situada en el hemisferio derecho, está vinculada específicamente al reconocimiento de rostros. [55]
En rigor, el reconocimiento es un proceso de recuperación de la memoria . Pero, en primer lugar, la forma en que se forma un recuerdo afecta la forma en que se recupera. Un área interesante de estudio relacionada con la memoria de reconocimiento trata sobre cómo los recuerdos se aprenden o codifican inicialmente en el cerebro. Este proceso de codificación es un aspecto importante de la memoria de reconocimiento porque determina no sólo si se reconoce o no un elemento introducido previamente, sino también cómo se recupera ese elemento a través de la memoria. Dependiendo de la fuerza de la memoria, el elemento puede ser "recordado" (es decir, un juicio de recuerdo) o simplemente "conocido" (es decir, un juicio de familiaridad). Por supuesto, la fuerza de la memoria depende de muchos factores, incluido si la persona prestó toda su atención a memorizar la información o si estaba distraída, si está intentando aprender activamente (aprendizaje intencional) o solo aprende pasivamente. si se les permitió ensayar la información o no, etc., aunque estos detalles contextuales están fuera del alcance de esta entrada.
Varios estudios han demostrado que cuando un individuo dedica toda su atención al proceso de memorización, la fuerza del recuerdo exitoso está relacionada con la magnitud de la activación bilateral en la corteza prefrontal, el hipocampo y la circunvolución parahipocampal. [56] [57] [58] Cuanto mayor sea la activación en estas áreas durante el aprendizaje, mejor será la memoria. Por lo tanto, estas áreas están involucradas en la formación de memorias recordativas detalladas. [59] Por el contrario, cuando los sujetos se distraen durante el proceso de codificación de la memoria, solo se activan la corteza prefrontal derecha y la circunvolución parahipocampal izquierda. [52] Estas regiones están asociadas con "una sensación de conocimiento" o familiaridad. [59] Dado que las áreas involucradas en la familiaridad también están involucradas en el recuerdo, esto se ajusta a una teoría de reconocimiento de proceso único, al menos en lo que respecta a la codificación de los recuerdos.
La memoria de reconocimiento no se limita al dominio visual; podemos reconocer cosas en cada una de las cinco modalidades sensoriales tradicionales (es decir, vista, oído, tacto, olfato y gusto). Aunque la mayoría de las investigaciones neurocientíficas se han centrado en el reconocimiento visual, también se han realizado estudios relacionados con la audición (oído), el olfato (olfato), el gusto (gusto) y la tactilidad (tacto).
La memoria de reconocimiento auditivo depende principalmente del lóbulo temporal medial, como lo demuestran los estudios en pacientes lesionados y amnésicos. [60] Además, los estudios realizados en monos [61] y perros [62] han confirmado que las lesiones de la corteza perininal y entorrinal no afectan la memoria de reconocimiento auditivo como lo hacen en la visión. Es necesario realizar más investigaciones sobre el papel del hipocampo en la memoria de reconocimiento auditivo, ya que los estudios en pacientes lesionados sugieren que el hipocampo desempeña un papel pequeño en la memoria de reconocimiento auditivo [60] , mientras que los estudios con perros lesionados [62] contradicen directamente este hallazgo. . También se ha propuesto que el área TH es vital para la memoria de reconocimiento auditivo [60] , pero también se deben realizar más investigaciones en esta área. Los estudios que comparan la memoria de reconocimiento visual y auditiva concluyen que la modalidad auditiva es inferior. [63]
La investigación sobre el olfato humano es escasa en comparación con otros sentidos como la visión y el oído, y los estudios dedicados específicamente al reconocimiento olfativo son aún más raros. Por tanto, la poca información que existe sobre este tema se obtiene a través de estudios en animales. Los roedores como los ratones o las ratas son sujetos adecuados para la investigación del reconocimiento de olores, dado que el olfato es su sentido principal. [64] "[Para estas especies], el reconocimiento de los olores corporales individuales es análogo al reconocimiento del rostro humano en el sentido de que proporciona información sobre la identidad". [65] En ratones, los olores corporales individuales están representados en el complejo mayor de histocompatibilidad (MHC). [65] En un estudio realizado con ratas, [66] se descubrió que la corteza orbitofrontal (OF) desempeña un papel importante en el reconocimiento de olores. La OF está conectada recíprocamente con las áreas perirrinal y entorrinal del lóbulo temporal medial, [66] que también han sido implicadas en la memoria de reconocimiento.
La memoria de reconocimiento gustativo, o reconocimiento del gusto, se correlaciona con la actividad en el lóbulo temporal anterior (ATL). [67] Además de las técnicas de imágenes cerebrales, el papel del ATL en el reconocimiento gustativo se evidencia por el hecho de que las lesiones en esta área dan como resultado un umbral aumentado para el reconocimiento del gusto en los humanos. [68] La neurotransmisión colinérgica en la corteza perirrinal es esencial para la adquisición de la memoria de reconocimiento del gusto y la aversión condicionada al gusto en humanos. [69]
Los monos con lesiones en las cortezas perirrinal y parahipocampal también muestran deterioro en el reconocimiento táctil. [70]
El concepto de especificidad de dominio ha ayudado a los investigadores a profundizar en los sustratos neuronales de la memoria de reconocimiento. La especificidad de dominio es la noción de que algunas áreas del cerebro son responsables casi exclusivamente del procesamiento de categorías particulares. Por ejemplo, está bien documentado que la circunvolución fusiforme (FFA) en el lóbulo temporal inferior está muy involucrada en el reconocimiento facial. Una región específica en esta circunvolución incluso se denomina área de la cara fusiforme debido a su mayor actividad neurológica durante la percepción de la cara . [71] De manera similar, también hay una región del cerebro conocida como área del lugar parahipocámpico en la circunvolución parahipocámpica . Como su nombre lo indica, esta área es sensible al contexto ambiental y a los lugares. [72] El daño a estas áreas del cerebro puede provocar déficits muy específicos. Por ejemplo, el daño a la FFA a menudo conduce a prosopagnosia , una incapacidad para reconocer rostros. [73] Las lesiones en varias regiones del cerebro como estas sirven como datos de estudios de casos que ayudan a los investigadores a comprender los correlatos neuronales del reconocimiento.

Los lóbulos temporales mediales y las estructuras que los rodean son de inmensa importancia para la memoria en general. El hipocampo es de particular interés. Está bien documentado que el daño aquí puede resultar en amnesia retrógrada o anterógrada severa, el paciente es incapaz de recordar ciertos eventos de su pasado o crear nuevos recuerdos, respectivamente. [74] Sin embargo, el hipocampo no parece ser el "depósito" de la memoria. Más bien, podría funcionar más bien como una estación repetidora. Las investigaciones sugieren que es a través del hipocampo que la memoria a corto plazo participa en el proceso de consolidación (la transferencia al almacenamiento a largo plazo). Los recuerdos se transfieren desde el hipocampo a la neocorteza lateral más amplia a través de la corteza entorrinal. [75] Esto ayuda a explicar por qué muchos amnésicos han preservado las capacidades cognitivas. Es posible que tengan una memoria normal a corto plazo, pero no pueden consolidarla y se pierden rápidamente. Las lesiones en el lóbulo temporal medial a menudo dejan al sujeto con la capacidad de aprender nuevas habilidades, también conocidas como memoria procedimental . Si experimenta amnesia anterógrada, el sujeto no puede recordar ninguna de las pruebas de aprendizaje, pero mejora constantemente con cada prueba. [76] Esto resalta el carácter distintivo del reconocimiento como un tipo de memoria particular y separado, que cae en el dominio de la memoria declarativa .
El hipocampo también es útil en la distinción entre familiaridad y recuerdo en el reconocimiento, como se mencionó anteriormente. Un recuerdo familiar es un recuerdo libre de contexto en el que la persona tiene una sensación de "saber", como en "Sé que puse las llaves de mi auto aquí en alguna parte". A veces se puede comparar con una sensación en la punta de la lengua. El recuerdo, por otro lado, es un proceso mucho más específico, deliberado y consciente, también denominado recuerdo. [4] Se cree que el hipocampo está muy involucrado en el recuerdo, mientras que la familiaridad se atribuye a la corteza perirrinal y a la corteza temporal más amplia en general; sin embargo, existe un debate sobre la validez de estos sustratos neuronales e incluso la separación entre familiaridad y recuerdo en sí. [77]
El daño a los lóbulos temporales también puede resultar en agnosia visual , un déficit en el que los pacientes son incapaces de reconocer objetos adecuadamente, ya sea debido a un déficit perceptivo o a un déficit en la memoria semántica. [78] En el proceso de reconocimiento de objetos, la información visual de los lóbulos occipitales (como líneas, movimiento, color, etc.) en algún momento debe ser interpretada activamente por el cerebro y atribuirle significado. Esto se conoce comúnmente en términos de vía ventral o "qué", que conduce a los lóbulos temporales. [79] Las personas con agnosia visual a menudo son capaces de identificar las características de un objeto (es pequeño, cilíndrico, tiene un mango, etc.), pero no pueden reconocer el objeto en su totalidad (una taza de té). [80] Esto se ha denominado específicamente agnosia integrativa . [78]
Durante mucho tiempo se pensó que la memoria de reconocimiento involucraba sólo las estructuras del lóbulo temporal medial . Investigaciones más recientes sobre neuroimagen han comenzado a demostrar que el lóbulo parietal también desempeña un papel importante, aunque a menudo sutil [81] , en la memoria de reconocimiento. Los primeros estudios de PET y fMRI demostraron la activación de la corteza parietal posterior durante las tareas de reconocimiento, [82] sin embargo, esto se atribuyó inicialmente a la activación de recuperación del precuneus , que se pensaba involucrado en el restablecimiento del contenido visual en la memoria. [83]
Nueva evidencia de estudios de pacientes con daño del lóbulo parietal posterior derecho indica déficits de reconocimiento muy específicos. [84] Este daño provoca un deterioro del rendimiento en tareas de reconocimiento de objetos con una variedad de estímulos visuales, incluidos colores, objetos familiares y formas nuevas. Este déficit de rendimiento no es el resultado de errores de seguimiento de la fuente , y el rendimiento preciso en las tareas de recuperación indica que la información ha sido codificada. Por lo tanto, el daño al lóbulo parietal posterior no causa errores globales de recuperación de la memoria, sólo errores en las tareas de reconocimiento.
El daño de la corteza parietal lateral (ya sea dextral o sinistral ) afecta el desempeño en tareas de memoria de reconocimiento, pero no afecta los recuerdos fuente. [85] Es más probable que lo que se recuerda sea del tipo "familiar" o "saber", en lugar de "recordar" o "recordar", [81] lo que indica que el daño a la corteza parietal perjudica la experiencia consciente de la memoria.
Existen varias hipótesis que buscan explicar la implicación del lóbulo parietal posterior en la memoria de reconocimiento. El modelo de atención a la memoria (AtoM) postula que el lóbulo parietal posterior podría desempeñar el mismo papel en la memoria que en la atención: mediar procesos de arriba hacia abajo versus procesos de abajo hacia arriba . [81] Los objetivos de la memoria pueden ser deliberados (de arriba hacia abajo) o en respuesta a una señal de memoria externa (de abajo hacia arriba). El lóbulo parietal superior sustenta objetivos de arriba hacia abajo, aquellos proporcionados por instrucciones explícitas. El lóbulo parietal inferior puede hacer que el lóbulo parietal superior redirija la atención a la memoria impulsada de abajo hacia arriba en presencia de una señal ambiental. Éste es el proceso de memoria espontáneo y no deliberado involucrado en el reconocimiento. Esta hipótesis explica muchos hallazgos relacionados con la memoria episódica, pero no explica el hallazgo de que la disminución de las señales de memoria de arriba hacia abajo dadas a pacientes con daño bilateral del lóbulo parietal posterior tuvo poco efecto sobre el rendimiento de la memoria. [86]
Una nueva hipótesis explica una mayor variedad de hallazgos de lesiones del lóbulo parietal al proponer que el papel del lóbulo parietal está en la experiencia subjetiva de viveza y confianza en los recuerdos. [81] Esta hipótesis está respaldada por hallazgos de que las lesiones en el lóbulo parietal causan la percepción de que los recuerdos carecen de viveza y dan a los pacientes la sensación de que su confianza en sus recuerdos está comprometida. [87]
La hipótesis del amortiguador de salida de la corteza parietal postula que las regiones parietales ayudan a retener el contenido cualitativo de los recuerdos para su recuperación y hacerlos accesibles a los procesos de toma de decisiones . [81] El contenido cualitativo de los recuerdos ayuda a distinguir los recordados, por lo que el deterioro de esta función reduce la confianza en los juicios de reconocimiento, como en los pacientes con lesiones del lóbulo parietal.
Varias otras hipótesis intentan explicar el papel del lóbulo parietal en la memoria de reconocimiento. La hipótesis del acumulador mnemotécnico postula que el lóbulo parietal contiene una señal de intensidad de la memoria, que se compara con criterios internos para realizar juicios de reconocimiento antiguos/nuevos. [81] Esto se relaciona con la teoría de detección de señales y explica que los elementos recordados se perciban como "más antiguos" que los elementos familiares. La hipótesis de la atención a la representación interna postula que las regiones parietales cambian y mantienen la atención a las representaciones de la memoria. [81] Esta hipótesis se relaciona con el modelo AtoM y sugiere que las regiones parietales están involucradas en una intención deliberada y de arriba hacia abajo de recordar.
Un posible mecanismo de la participación del lóbulo parietal en la memoria de reconocimiento puede ser la activación diferencial de los recuerdos recordados frente a los familiares, y de los estímulos antiguos frente a los nuevos. Esta región del cerebro muestra una mayor activación durante segmentos de tareas de reconocimiento que contienen principalmente estímulos antiguos, frente a estímulos principalmente nuevos. [82] Se ha demostrado una disociación entre las regiones parietales dorsal y ventral , donde la región ventral experimenta más activación para los elementos recordados y la región dorsal experimenta más activación para los elementos familiares. [81]
La anatomía proporciona más pistas sobre el papel del lóbulo parietal en la memoria de reconocimiento. La corteza parietal lateral comparte conexiones con varias regiones del lóbulo temporal medial , incluidas sus regiones hipocámpica , parahipocámpica y entorrinal . [81] Estas conexiones pueden facilitar la influencia del lóbulo temporal medial en el procesamiento de información cortical. [82]
La evidencia de pacientes amnésicos ha demostrado que las lesiones en el lóbulo frontal derecho son una causa directa de errores de reconocimiento falso. Algunos sugieren que esto se debe a una variedad de factores que incluyen procesos defectuosos de monitoreo, recuperación y decisión. [88] Los pacientes con lesiones del lóbulo frontal también mostraron evidencia de un marcado deterioro de la memoria facial anterógrada y relativamente leve retrógrada. [89]
La capacidad de reconocer estímulos como viejos o nuevos tiene importantes ventajas evolutivas para los humanos. Discernir entre estímulos familiares y desconocidos permite evaluar rápidamente las amenazas en entornos a menudo hostiles. La velocidad y la precisión de un juicio de reconocimiento antiguo/nuevo son dos componentes de una serie de procesos cognitivos que permiten a los humanos identificar y responder a peligros potenciales en su entorno. [90] El reconocimiento de un suceso anterior es una adaptación que proporciona una pista de la utilidad de la información para los procesos de toma de decisiones . [90]
La corteza perirrinal participa notablemente tanto en la respuesta al miedo como en la memoria de reconocimiento. [91] Las neuronas en esta región se activan fuertemente en respuesta a nuevos estímulos y se activan con menos frecuencia a medida que aumenta la familiaridad con el estímulo. [17] La información sobre la identidad del estímulo llega al hipocampo a través de la corteza perirrinal , [92] y el sistema perirrinal contribuye con una evaluación rápida y automática de la familiaridad de los estímulos y lo reciente de su presentación. [93] Esta respuesta de reconocimiento tiene la clara ventaja evolutiva de proporcionar información para los procesos de toma de decisiones de una manera automatizada, conveniente y sin esfuerzo, lo que permite respuestas más rápidas a las amenazas.
Una aplicación práctica de la memoria de reconocimiento es en relación con el desarrollo de pruebas de opción múltiple en un entorno académico. Una buena prueba no recurre a la memoria de reconocimiento, sino que quiere discernir qué tan bien una persona codificó y puede recordar un concepto. Si las personas confían en el reconocimiento para utilizarlo en una prueba de memoria (como la de opción múltiple), pueden reconocer una de las opciones, pero esto no significa necesariamente que sea la respuesta correcta. [94]
{{cite journal}}: Mantenimiento CS1: varios nombres: lista de autores ( enlace )