La ficoeritrina ( PE ) es un complejo de proteína- pigmento rojo de la familia de las ficobiliproteínas captadoras de luz , presente en cianobacterias , [1] algas rojas [2] y criptofitas , [3] accesorio a los principales pigmentos de clorofila responsables de la fotosíntesis . El pigmento rojo se debe al grupo prostético, ficoeritrobilina, que le da a la ficoeritrina su color rojo. [4]
Como todas las ficobiliproteínas, está compuesta por una parte proteica que se une covalentemente a cromóforos llamados ficobilinas . En la familia de las ficoeritrinas, las ficobilinas más conocidas son: ficoeritrobilina , el cromóforo aceptor típico de ficoeritrina. La ficoeritrobilina es una molécula tetrapirrol lineal que se encuentra en cianobacterias, algas rojas y criptomonas. Junto con otras bilinas como la ficocianobilina, sirve como pigmento recolector de luz en las estructuras fotosintéticas recolectoras de luz de las cianobacterias llamadas ficobilisomas. [5] Las ficoeritrinas están compuestas de monómeros (αβ), generalmente organizados en un trímero en forma de disco (αβ) 3 o hexámero (αβ) 6 (el segundo es la unidad funcional de las varillas de antena ). Estos complejos típicos también contienen un tercer tipo de subunidad, la cadena γ. [2]

Las ficobiliproteínas forman parte de enormes complejos proteicos de antenas que captan luz, llamados ficobilisomas . En las algas rojas, están ancladas al lado estromal de las membranas tilacoides de los cloroplastos , mientras que en las criptofitas, los ficobilisomas están reducidos y (moléculas de ficobiliproteína 545 PE545 aquí) están densamente empaquetados dentro del lumen de los tilacoides. [3] [7]
Las ficobiliproteínas tienen muchas aplicaciones prácticas, incluidas propiedades importantes como la actividad hepatoprotectora, antioxidante, antiinflamatoria y antienvejecimiento de las PBP, lo que permite su uso en las industrias alimentaria, cosmética, farmacéutica y biomédica. También se ha observado que las PBP muestran un efecto beneficioso en la terapéutica de algunas enfermedades como el Alzheimer y el cáncer. [8]
La ficoeritrina es un pigmento accesorio de los principales pigmentos clorofílicos responsables de la fotosíntesis . La energía luminosa es captada por la ficoeritrina y luego se transmite al par de clorofilas del centro de reacción, la mayoría de las veces a través de las ficobiliproteínas ficocianina y aloficocianina .
Las ficoeritrinas, excepto la ficoeritrina 545 (PE545), están compuestas de monómeros (αβ) ensamblados en 6 hexámeros (αβ) o 3 trímeros (αβ) con forma de disco con simetría 32 o 3 y que encierran un canal central. En los ficobilisomas (PBS), cada trímero o hexámero contiene al menos una proteína de enlace ubicada en el canal central. La B-ficoeritrina (B-PE) y la R-ficoeritrina (R-PE) de las algas rojas , además de las cadenas α y β, tienen una tercera subunidad γ que contribuye tanto con funciones de enlace como de captación de luz, porque contiene cromóforos. [2]
La R-ficoeritrina es producida predominantemente por algas rojas. La proteína está formada por al menos tres subunidades diferentes y varía según la especie de alga que la produce. La estructura de subunidad de la R-PE más común es (αβ) 6 γ. La subunidad α tiene dos ficoeritrobilinas (PEB), la subunidad β tiene 2 o 3 PEB y una ficourobilina (PUB), mientras que se informa que las diferentes subunidades gamma tienen 3 PEB y 2 PUB (γ 1 ) o 1 o 2 PEB y 1 PUB (γ 2 ). El peso molecular de la R-PE es de 250.000 daltons.
Las estructuras cristalinas disponibles en el Protein Data Bank [12] contienen en una unidad asimétrica (αβ) 2 o (αβγ) 2 diferentes ficoeritrinas:
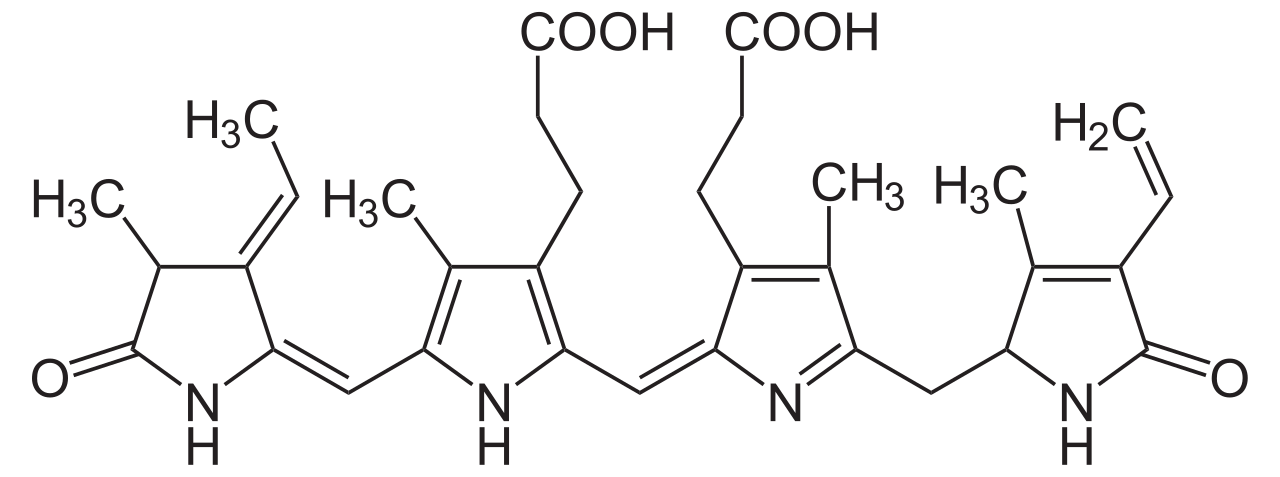

La molécula biológica supuesta de la ficoeritrina 545 (PE545) es (αβ) 2 o más bien (α 3 β)(α 2 β) . Los números 2 y 3 después de las letras α en la segunda fórmula son parte de los nombres de las cadenas, no sus conteos. El nombre criptofitano sinónimo de la cadena α 3 es cadena α 1 .
El conjunto más grande de B-ficoeritrina (B-PE) es el trímero (αβ) 3 [9] [10] . Sin embargo, las preparaciones a partir de algas rojas también producen el hexámero (αβ) 6 [2] . En el caso de R-ficoeritrina (R-PE) la molécula biológica más grande asumida aquí es (αβγ) 6 , (αβγ) 3 (αβ) 3 o (αβ) 6 dependiendo de la publicación, para otros tipos de ficoeritrina (αβ) 6 . Estas cadenas γ del Protein Data Bank son muy pequeñas y constan solo de tres o seis aminoácidos reconocibles [15] [16] , mientras que la cadena γ del enlazador descrita al comienzo de esta sección es grande (por ejemplo, 277 aminoácidos de longitud 33 kDa en el caso de γ 33 del alga roja Aglaothamnion neglectum ) [17] [2] . Esto se debe a que la densidad electrónica del polipéptido gamma se promedia principalmente por su simetría cristalográfica triple y solo se pueden modelar unos pocos aminoácidos [15] [16] [18] [19] .
Para (αβγ) 6 , (αβ) 6 o (αβγ) 3 (αβ) 3 los valores de la tabla deben simplemente multiplicarse por 3, (αβ) 3 contiene cantidades intermedias de moléculas no proteicas. (esta incongruencia debe corregirse)
En la ficoeritrina PE545 anterior, una cadena α (-2 o -3) se une a una molécula de billina; en otros ejemplos, se une a dos moléculas. La cadena β siempre se une a tres moléculas. La pequeña cadena γ no se une a ninguna.
Dos moléculas de N -metil asparagina están unidas a la cadena β, una 5-hidroxilisina a α (-3 o -2), una Mg 2+ a α-3 y β, una Cl − a β, 1–2 moléculas de SO2−
4a α o β.
A continuación se muestra un ejemplo de la estructura cristalina de la R-ficoeritrina del Protein Data Bank :

Los picos de absorción en el espectro de luz visible se miden a 495 y 545/566 nm, dependiendo de los cromóforos unidos y del organismo considerado. Existe un pico de emisión fuerte a 575 ± 10 nm. (La ficoeritrina absorbe una luz ligeramente azul-verde/amarillenta y emite una luz ligeramente naranja-amarillenta).
Las bilinas PEB y DBV en PE545 también absorben en la región espectral verde, con máximos a 545 y 569 nm respectivamente. El máximo de emisión de fluorescencia se encuentra a 580 nm. [3]

Como se mencionó anteriormente, la ficoeritrina se puede encontrar en una variedad de especies de algas. Por lo tanto, puede haber variación en la eficiencia de la absorción y emisión de luz requerida para facilitar la fotosíntesis. Esto podría ser el resultado de la profundidad en la columna de agua en la que reside típicamente un alga específica y la consiguiente necesidad de una mayor o menor eficiencia de los pigmentos accesorios.
Gracias a los avances en la tecnología de detección e imagen que pueden evitar el rápido fotoblanqueo, los fluoróforos proteicos se han convertido en una herramienta viable y poderosa para los investigadores en campos como la microscopía, el análisis de microarrays y el Western blotting. En vista de esto, puede ser beneficioso para los investigadores examinar estas R-ficoeritrinas variables para determinar cuál es la más apropiada para su aplicación particular. Incluso un pequeño aumento en la eficiencia de la fluorescencia podría reducir el ruido de fondo y disminuir la tasa de resultados falsos negativos.
La R-ficoeritrina (también conocida como PE o R-PE) es útil en el laboratorio como un indicador basado en fluorescencia para la presencia de cianobacterias y para marcar anticuerpos , más a menudo para citometría de flujo . También se utiliza en ensayos de microarrays, ELISA y otras aplicaciones que requieren alta sensibilidad pero no fotoestabilidad. [20] Su uso es limitado en microscopía de inmunofluorescencia debido a sus características de fotoblanqueo rápido . También hay otros tipos de ficoeritrinas, como la B-ficoeritrina, que tienen propiedades espectrales ligeramente diferentes. La B-ficoeritrina absorbe fuertemente a aproximadamente 545 nm (verde ligeramente amarillento) y emite fuertemente a 572 nm (amarillo) en cambio y podría ser más adecuada para algunos instrumentos. La B-ficoeritrina también puede ser menos "pegajosa" que la R-ficoeritrina y contribuye menos a la señal de fondo debido a la unión no específica en ciertas aplicaciones. [ cita requerida ] Sin embargo, el R-PE está mucho más disponible como conjugado de anticuerpo. Ficoeritrina (PE, λA máx. = 540–570 nm; λF máx. = 575–590 nm) [8]
La R-ficoeritrina y la B-ficoeritrina se encuentran entre los colorantes fluorescentes más brillantes jamás identificados.
{{cite journal}}: Requiere citar revista |journal=( ayuda ){{cite journal}}: Requiere citar revista |journal=( ayuda ){{cite journal}}: Requiere citar revista |journal=( ayuda ){{cite journal}}: Requiere citar revista |journal=( ayuda )