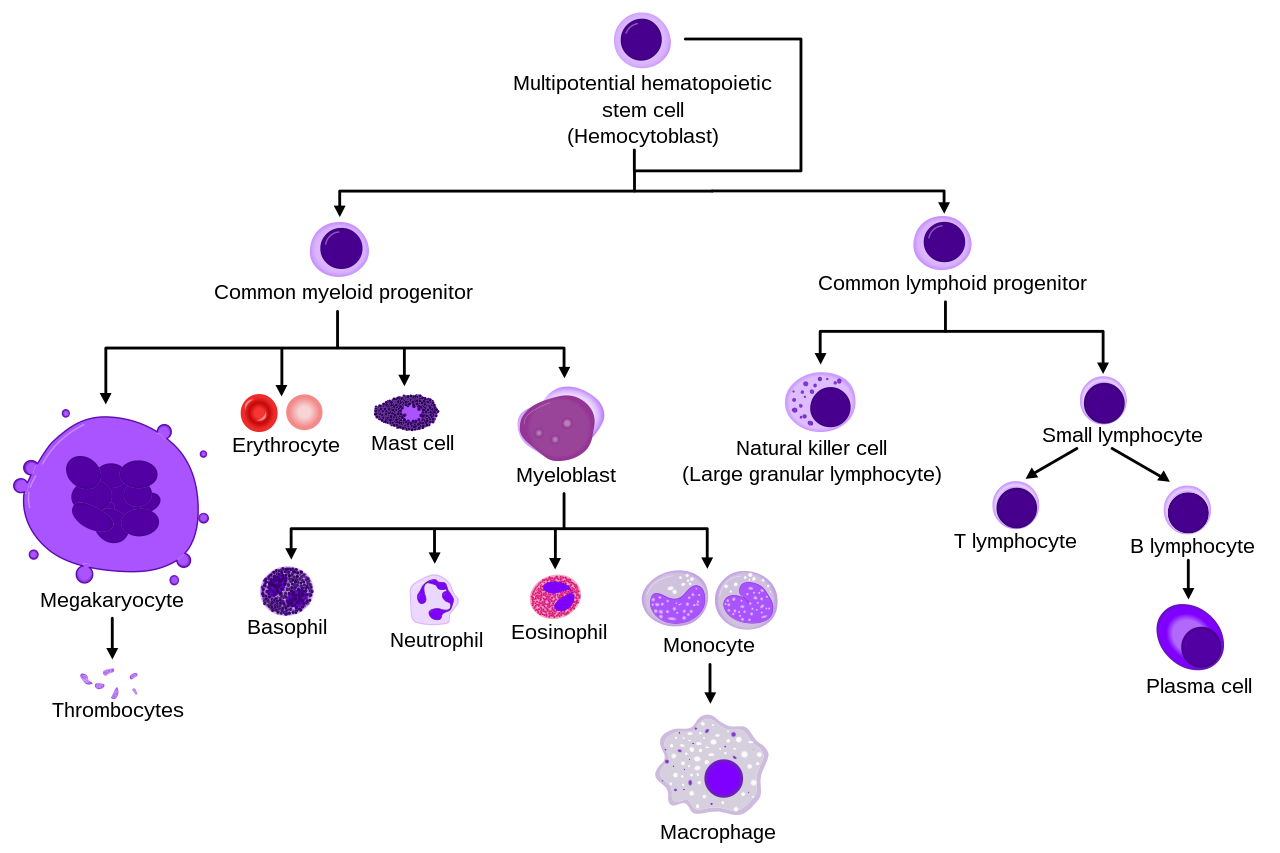
Diagrama que muestra el desarrollo de diferentes células sanguíneas, desde células madre hematopoyéticas hasta células maduras.
Hematopoyesis ( / h ɪ ˌ m æ t ə p ɔɪ ˈ iː s ɪ s , ˌ h iː m ə t oʊ - , ˌ h ɛ m ə -/ , [1] [2] del griego αἷμα , 'sangre' y ποιεῖν 'to make'; también hematopoyesis en inglés americano; a veces también h(a)emopoyesis ) es la formación de componentes celulares sanguíneos . Todos los componentes sanguíneos celulares se derivan de células madre hematopoyéticas . [3] En un ser humano adulto sano, aproximadamente diez mil millones (1010 ) a cien mil millones (1011 ) se producen nuevas células sanguíneas por día, con el fin de mantener niveles estables en la circulación periférica. [4] [5] [ página necesaria ]
Proceso
Células madre hematopoyéticas (HSC)
Las células madre hematopoyéticas (HSC) residen en la médula ósea ( médula ósea ) y tienen la capacidad única de dar lugar a todos los diferentes tipos y tejidos de células sanguíneas maduras. [3] Las HSC son células que se renuevan a sí mismas: cuando se diferencian, al menos algunas de sus células hijas permanecen como HSC para que el conjunto de células madre no se agote. [6] Este fenómeno se llama división asimétrica. [7] Las otras hijas de HSC ( células progenitoras mieloides y linfoides ) pueden seguir cualquiera de las otras vías de diferenciación que conducen a la producción de uno o más tipos específicos de células sanguíneas, pero no pueden renovarse. El grupo de progenitores es heterogéneo y se puede dividir en dos grupos; HSC autorrenovables a largo plazo y HSC autorrenovables solo de forma transitoria, también llamados de corto plazo. [8] Este es uno de los principales procesos vitales del cuerpo.
tipos de células
Todas las células sanguíneas se dividen en tres linajes. [9]
Los glóbulos rojos , que también se llaman eritrocitos, son las células transportadoras de oxígeno . Los eritrocitos son funcionales y se liberan a la sangre. El número de reticulocitos, que son glóbulos rojos inmaduros, da una estimación de la tasa de eritropoyesis .
La granulopoyesis (o granulocitopoyesis) es la hematopoyesis de los granulocitos, excepto los mastocitos que son granulocitos pero con una maduración extramedular. [10]
Entre 1948 y 1950, el Comité para la Aclaración de la Nomenclatura de las Células y las Enfermedades de la Sangre y de los Órganos Sanguíneos publicó informes sobre la nomenclatura de las células sanguíneas. [11] [12] A continuación se muestra una descripción general de la terminología, desde la etapa de desarrollo más temprana hasta la final:
[raíz] explosión
pro[raíz]cito
[raíz]cito
meta[raíz]cito
nombre de celular maduro
La raíz de unidades formadoras de colonias de eritrocitos (UFC-E) es "rubri", de unidades formadoras de colonias de granulocitos-monocitos (UFC-GM) es "granulo" o "mielo" y "mono", de unidades formadoras de colonias de linfocitos. (CFU-L) es "linfo" y para unidades formadoras de colonias de megacariocitos (CFU-Meg) es "megacario". Según esta terminología, las etapas de formación de los glóbulos rojos serían: rubriblasto, prorubricito, rubricito, metarubricito y eritrocito. Sin embargo, la siguiente nomenclatura parece ser, en la actualidad, la más frecuente:
Los osteoclastos también surgen de células hematopoyéticas del linaje monocitos/neutrófilos, específicamente CFU-GM.
Ubicación
Sitios de hematopoyesis (humana) en períodos pre y posnatales.
En los embriones en desarrollo, la formación de sangre se produce en agregados de células sanguíneas en el saco vitelino, llamados islas de sangre . A medida que avanza el desarrollo, se produce formación de sangre en el bazo , el hígado y los ganglios linfáticos . [13] Cuando la médula ósea se desarrolla, eventualmente asume la tarea de formar la mayoría de las células sanguíneas de todo el organismo. [3] Sin embargo, la maduración, activación y cierta proliferación de células linfoides ocurre en el bazo, el timo y los ganglios linfáticos. En los niños, la hematopoyesis se produce en la médula de los huesos largos como el fémur y la tibia. En los adultos, ocurre principalmente en la pelvis, el cráneo, las vértebras y el esternón. [14]
extramedular
En algunos casos, el hígado, el timo y el bazo pueden reanudar su función hematopoyética, si es necesario. Esto se llama hematopoyesis extramedular . Puede hacer que estos órganos aumenten sustancialmente de tamaño. Durante el desarrollo fetal, dado que los huesos y, por tanto, la médula ósea se desarrollan más tarde, el hígado funciona como el principal órgano hematopoyético. Por tanto, el hígado aumenta de tamaño durante el desarrollo. [15] La hematopoyesis y la mielopoyesis extramedulares pueden suministrar leucocitos en enfermedades cardiovasculares e inflamación durante la edad adulta. [16] [17] Los macrófagos esplénicos y las moléculas de adhesión pueden estar involucrados en la regulación de la generación de células mieloides extramedulares en la enfermedad cardiovascular . [18] [19]
Maduración
Diagrama más detallado y completo que muestra el desarrollo de diferentes células sanguíneas en humanos.
Las características morfológicas de las células hematopoyéticas se muestran como se ve en una tinción de Wright, tinción de May-Giemsa o tinción de May-Grünwald-Giemsa. Los nombres alternativos de determinadas celdas se indican entre paréntesis.
Ciertas células pueden tener más de una apariencia característica. En estos casos se ha incluido más de una representación de una misma celda.
Juntos, los monocitos y los linfocitos comprenden los agranulocitos, a diferencia de los granulocitos (basófilos, neutrófilos y eosinófilos) que se producen durante la granulopoyesis.
B., N. y E. representan basófilos, neutrofílicos y eosinófilos, respectivamente, como en promielocitos basófilos. Para los linfocitos, T y B son designaciones reales.
El eritrocito policromático (reticulocito) de la derecha muestra su aspecto característico cuando se tiñe con azul de metileno o Azure B.
El eritrocito de la derecha es una representación más precisa de su apariencia real cuando se ve a través de un microscopio.
Otras células que surgen del monocito: osteoclastos, microglía (sistema nervioso central), células de Langerhans (epidermis), células de Kupffer (hígado).
Para mayor claridad, los linfocitos T y B se dividen para indicar mejor que la célula plasmática surge de la célula B. Tenga en cuenta que no hay diferencia en la apariencia de las células B y T a menos que se aplique una tinción específica.
A medida que una célula madre madura, sufre cambios en la expresión genética que limitan los tipos de células en los que puede convertirse y la acercan a un tipo de célula específico ( diferenciación celular ). Estos cambios a menudo pueden rastrearse monitoreando la presencia de proteínas en la superficie de la célula. Cada cambio sucesivo acerca la célula al tipo celular final y limita aún más su potencial para convertirse en un tipo celular diferente. [ cita necesaria ]
Determinación del destino celular
Se han propuesto dos modelos de hematopoyesis: el determinismo y la teoría estocástica. [20] Para las células madre y otras células sanguíneas indiferenciadas en la médula ósea, la determinación generalmente se explica por la teoría del determinismo de la hematopoyesis, diciendo que los factores estimulantes de colonias y otros factores del microambiente hematopoyético determinan que las células sigan un determinado camino de diferenciación celular. [3] Esta es la forma clásica de describir la hematopoyesis. En la teoría estocástica , las células sanguíneas indiferenciadas se diferencian en tipos de células específicos de forma aleatoria. Esta teoría ha sido respaldada por experimentos que muestran que dentro de una población de células progenitoras hematopoyéticas de ratón, la variabilidad estocástica subyacente en la distribución de Sca-1 , un factor de células madre , subdivide la población en grupos que exhiben tasas variables de diferenciación celular . Por ejemplo, bajo la influencia de la eritropoyetina (un factor de diferenciación de eritrocitos), una subpoblación de células (definida por los niveles de Sca-1) se diferenciaron en eritrocitos a un ritmo siete veces mayor que el resto de la población. [21] Además, se demostró que si se le permitía crecer, esta subpoblación restablecía la subpoblación original de células, lo que respalda la teoría de que se trata de un proceso estocástico y reversible. Otro nivel en el que la estocasticidad puede ser importante es en el proceso de apoptosis y autorrenovación. En este caso, el microambiente hematopoyético prevalece para que algunas células sobrevivan y otras, en cambio, realicen apoptosis y mueran. [3] Al regular este equilibrio entre diferentes tipos de células, la médula ósea puede alterar la cantidad de diferentes células que finalmente se producirán. [22]
La producción de glóbulos rojos y blancos está regulada con gran precisión en humanos sanos y la producción de leucocitos aumenta rápidamente durante la infección. La proliferación y autorrenovación de estas células depende de factores de crecimiento. Uno de los actores clave en la autorrenovación y el desarrollo de las células hematopoyéticas es el factor de células madre (SCF), [25] que se une al receptor c-kit en las HSC. La ausencia de SCF es letal. Existen otros factores de crecimiento de glicoproteínas importantes que regulan la proliferación y maduración, como las interleucinas IL-2 , IL-3 , IL-6 , IL-7 . Otros factores, denominados factores estimulantes de colonias (CSF, por sus siglas en inglés), estimulan específicamente la producción de células comprometidas. Tres LCR son el LCR de granulocitos y macrófagos (GM-CSF), el LCR de granulocitos (G-CSF) y el LCR de macrófagos (M-CSF). [26] Estos estimulan la formación de granulocitos y son activos en las células progenitoras o en las células producto finales.
La eritropoyetina es necesaria para que una célula progenitora mieloide se convierta en eritrocito. [23] Por otro lado, la trombopoyetina hace que las células progenitoras mieloides se diferencien en megacariocitos ( células formadoras de trombocitos ). [23] El diagrama de la derecha proporciona ejemplos de citocinas y las células sanguíneas diferenciadas a las que dan lugar. [27]
Factores de transcripción
Los factores de crecimiento inician vías de transducción de señales , que conducen a la activación de factores de transcripción . Los factores de crecimiento provocan diferentes resultados según la combinación de factores y la etapa de diferenciación de la célula. Por ejemplo, la expresión a largo plazo de PU.1 da como resultado un compromiso mieloide y la inducción a corto plazo de la actividad de PU.1 conduce a la formación de eosinófilos inmaduros. [28] Recientemente, se informó que los factores de transcripción como NF-κB pueden regularse mediante microARN (p. ej., miR-125b) en la hematopoyesis. [29]
El primer actor clave en la diferenciación de HSC a un progenitor multipotente (MPP) es la proteína de unión α potenciadora del factor de transcripción CCAAT ( C/EBP α). "Las mutaciones en C/EBPα están asociadas con la leucemia mieloide aguda ". [30] A partir de este punto, las células pueden diferenciarse a lo largo del linaje eritroide-megacariocito o del linaje linfoide y mieloide, que tienen un progenitor común, llamado progenitor multipotente preparado con linfoide. Hay dos factores de transcripción principales. PU.1 para el linaje eritroide-megacariocito y GATA-1 , que conduce a un progenitor multipotente preparado con linfoides. [31]
Otros factores de transcripción incluyen Ikaros [32] ( desarrollo de células B ) y Gfi1 [33] (promueve el desarrollo de Th2 e inhibe Th1) o IRF8 [34] ( basófilos y mastocitos ). Es significativo que ciertos factores provocan diferentes respuestas en diferentes etapas de la hematopoyesis. Por ejemplo, CEBPα en el desarrollo de neutrófilos o PU.1 en monocitos y desarrollo de células dendríticas. Es importante señalar que los procesos no son unidireccionales: las células diferenciadas pueden recuperar atributos de las células progenitoras. [1]
Un ejemplo es el factor PAX5 , que es importante en el desarrollo de las células B y está asociado con los linfomas. [35] Sorprendentemente, los ratones con desactivación condicional de pax5 permitieron que las células B maduras periféricas se desdiferenciaran a progenitores tempranos de la médula ósea. Estos hallazgos muestran que los factores de transcripción actúan como cuidadores del nivel de diferenciación y no sólo como iniciadores. [36]
Las mutaciones en los factores de transcripción están estrechamente relacionadas con los cánceres de la sangre, como la leucemia mieloide aguda (AML) o la leucemia linfoblástica aguda (ALL). Por ejemplo, se sabe que Ikaros es regulador de numerosos eventos biológicos. Los ratones sin Ikaros carecen de células B , asesinas naturales y células T. [37] Ikaros tiene seis dominios de dedos de zinc , cuatro son dominios de unión al ADN conservados y dos son para dimerización . [38] Un hallazgo muy importante es que diferentes dedos de zinc participan en la unión a diferentes lugares del ADN y esta es la razón del efecto pleiotrópico de Ikaros y su diferente participación en el cáncer, pero principalmente son mutaciones asociadas con pacientes con BCR-Abl y es marcador de mal pronóstico. [39]
Otros animales
En algunos vertebrados , la hematopoyesis puede ocurrir en cualquier lugar donde haya un estroma laxo de tejido conectivo y un suministro sanguíneo lento, como el intestino , el bazo o el riñón . [40]
A diferencia de los mamíferos euterios, el hígado de los marsupiales recién nacidos es activamente hematopoyético. [41] [42] [43] [44]
^ "hematopoyesis". Dictionary.com íntegro (en línea). y nd . Consultado el 16 de octubre de 2019 .
^ abcde Birbrair, Alejandro; Frenette, Paul S. (1 de marzo de 2016). "Heterogeneidad de nicho en la médula ósea". Anales de la Academia de Ciencias de Nueva York . 1370 (1): 82–96. Código Bib : 2016NYASA1370...82B. doi :10.1111/nyas.13016. ISSN 1749-6632. PMC 4938003 . PMID 27015419.
^ Conferencias médicas del cuarto semestre en la Universidad de Uppsala 2008 por Leif Jansson
^ Monga I, Kaur K, Dhanda S (marzo de 2022). "Revisando la hematopoyesis: aplicaciones de la transcriptómica unicelular y masiva que analizan la heterogeneidad transcripcional en células madre hematopoyéticas". Sesiones informativas en genómica funcional . 21 (3): 159-176. doi :10.1093/bfgp/elac002. PMID 35265979.
^ Morrison, J.; Judith Kimble (2006). "Divisiones de células madre asimétricas y simétricas en el desarrollo y el cáncer" (PDF) . Naturaleza . 441 (7097): 1068–74. Código Bib : 2006Natur.441.1068M. doi : 10.1038/naturaleza04956. hdl : 2027.42/62868 . PMID 16810241. S2CID 715049.
^ Morrison SJ, Weissman IL (noviembre de 1994). "El subconjunto de células madre hematopoyéticas que se repobla a largo plazo es determinista y se puede aislar por fenotipo". Inmunidad . 1 (8): 661–73. doi :10.1016/1074-7613(94)90037-x. PMID 7541305.
^ "Hematopoyesis a partir de células madre pluripotentes". Biblioteca de recursos de anticuerpos . ThermoFisher científico . Consultado el 25 de abril de 2020 .
^ Mahler (2013). Haschek, Wanda; Rousseaux, Colin G.; Wallig, Matthew A. (eds.). Manual de patología toxicológica de Haschek y Rousseaux . editores asociados, Brad Bolon y Ricardo Ochoa; editora de ilustraciones, Beth W. (Tercera ed.). [Sl]: Prensa Académica. pag. 1863.ISBN978-0-12-415759-0.
↑ «PRIMER informe del comité para el esclarecimiento de la nomenclatura de células y enfermedades de la sangre y de los órganos hematopoyéticos» . Revista americana de patología clínica . 18 (5): 443–50. Mayo de 1948. doi :10.1093/ajcp/18.5_ts.443. PMID 18913573.
↑ «TERCER, cuarto y quinto informes del comité para el esclarecimiento de la nomenclatura de las células y enfermedades de la sangre y de los órganos hematopoyéticos» . Revista americana de patología clínica . 20 (6): 562–79. Junio de 1950. doi : 10.1093/ajcp/20.6.562. PMID 15432355.
^ Singh, Ranbir; Soman-Faulkner, Kristina; Sugumar, Kavin (2022). "Embriología, Hematopoyesis". NCBI . Estadísticas de perlas. PMID 31334965 . Consultado el 4 de septiembre de 2022 .
^ Fernández KS, de Alarcón PA (diciembre de 2013). "Desarrollo del sistema hematopoyético y trastornos de la hematopoyesis que se presentan durante la infancia y primera infancia" . Clínicas Pediátricas de América del Norte . 60 (6): 1273–89. doi :10.1016/j.pcl.2013.08.002. PMID 24237971.
^ Georgiades CS, Neyman EG, Francis IR, Sneider MB, Fishman EK (noviembre de 2002). "Presentaciones típicas y atípicas de la hematopoyesis extramedular". AJR. Revista Estadounidense de Roentgenología . 179 (5): 1239–43. doi : 10.2214/ajr.179.5.1791239 . PMID 12388506.
^ Swirski, Filip K.; Libby, Pedro; Aikawa, Elena; Alcaide, Pilar; Luscinskas, F. William; Weissleder, Ralph; Pittet, Mikael J. (2 de enero de 2007). "Los monocitos Ly-6Chi dominan la monocitosis asociada a hipercolesterolemia y dan lugar a macrófagos en los ateromas". Revista de investigación clínica . 117 (1): 195–205. doi : 10.1172/JCI29950 . PMC 1716211 . PMID 17200719.
^ Swirski FK, Nahrendorf M, Etzrodt M, Wildgruber M, Cortez-Retamozo V, Panizzi P, Figueiredo JL, Kohler RH, Chudnovskiy A, Waterman P, Aikawa E, Mempel TR, Libby P, Weissleder R, Pittet MJ (30 de julio) 2009). "Identificación de monocitos del reservorio esplénico y su implementación en sitios inflamatorios". Ciencia . 325 (5940): 612–616. Código Bib : 2009 Ciencia... 325..612S. doi : 10.1126/ciencia.1175202. PMC 2803111 . PMID 19644120.
^ Dutta, P; Hoyer, FF; Grigoryeva, LS; Sager, HB; Leuschner, F; Cortesanos, G; Borodovsky, A; Novobrantseva, T; Ruda, VM; Fitzgerald, K; Iwamoto, Y; Wojtkiewicz, G; Sol, Y; Da Silva, N; Libby, P; Anderson, director general; Swirski, FK; Weissleder, R; Nahrendorf, M (6 de abril de 2015). "Los macrófagos retienen células madre hematopoyéticas en el bazo a través de VCAM-1". La Revista de Medicina Experimental . 212 (4): 497–512. doi : 10.1084/jem.20141642 . PMC 4387283 . PMID 25800955.
^ Dutta, P; Hoyer, FF; Sol, Y; Iwamoto, Y; Punto, B; Weissleder, R; Magnani, JL; Swirski, FK; Nahrendorf, M (septiembre de 2016). "La inhibición de la E-selectina mitiga la activación de HSC esplénica y la mielopoyesis en ratones hipercolesterolémicos con infarto de miocardio". Arteriosclerosis, trombosis y biología vascular . 36 (9): 1802–8. doi : 10.1161/ATVBAHA.116.307519 . PMC 5001901 . PMID 27470513.
^ Kimmel, Marek (1 de enero de 2014). "Estocasticidad y determinismo en modelos de hematopoyesis". Un enfoque de biología de sistemas para la sangre . Avances en Medicina y Biología Experimentales. vol. 844, págs. 119-152. doi :10.1007/978-1-4939-2095-2_7. ISBN978-1-4939-2094-5. ISSN 0065-2598. PMID 25480640.
^ Chang, Hannah H.; Hemberg, Martín; Barahona, Mauricio; Ingber, Donald E.; Huang, Sui (2008). "El ruido de todo el transcriptoma controla la elección del linaje en células progenitoras de mamíferos". Naturaleza . 453 (7194): 544–547. Código Bib :2008Natur.453..544C. doi : 10.1038/naturaleza06965. PMC 5546414 . PMID 18497826.
^ Alenzi, FQ; Alenazi, BQ; Ahmad, SY; Salem, ML; Al-Jabri, AA; Wyse, RK (marzo de 2009). "La célula madre hematopoyética: entre la apoptosis y la autorrenovación". La Revista de Biología y Medicina de Yale . 82 (1): 7–18. PMC 2660591 . PMID 19325941.
^ abcd Biología celular molecular. Lodish, Harvey F. 5. ed. : – Nueva York: WH Freeman and Co., 2003, 973 s. factura. ISBN 0-7167-4366-3Lodish H, Berk A, Zipursky SL, Matsudaira P, Baltimore D, Darnell J (2000). "Los cánceres se originan en células en proliferación". Biología celular molecular (4ª ed.). Nueva York: WH Freeman. Figura 24-8: Formación de células sanguíneas diferenciadas a partir de células madre hematopoyéticas en la médula ósea. ISBN 0-7167-3136-3– a través de NCBI Bookshelf.
^ Flor de varilla; Humphrey P. sonó; Maureen M. Dale; Ritter, James M. (2007). Farmacología de Rang & Dale . Edimburgo: Churchill Livingstone. ISBN978-0-443-06911-6.
^ Broudy, VC (15 de agosto de 1997). "Factor de células madre y hematopoyesis". Sangre . 90 (4): 1345–64. doi : 10.1182/sangre.V90.4.1345 . PMID 9269751.
^ Ketley, Nueva Jersey; AC Newland (1997). "Factores de crecimiento hematopoyético". Postgrado Med J. 73 (858): 215–221. doi :10.1136/pgmj.73.858.215. PMC 2431295 . PMID 9156123.
^ Hauke, Ralph; Stefano R. Tarantolo (noviembre de 2000). "Factores de crecimiento hematopoyético". Medicina de laboratorio . 31 (11): 613–5. doi : 10.1309/HNTM-ELUV-AV9G-MA1P .
^ Engel, yo; Murre, C (octubre de 1999). "Factores de transcripción en la hematopoyesis" . Opinión actual en genética y desarrollo . 9 (5): 575–9. doi :10.1016/s0959-437x(99)00008-8. PMID 10508690.
^ O'Connell, R; Rao, D.; Baltimore, D (2012). "Regulación de microARN de respuestas inflamatorias". Revista Anual de Inmunología . 30 : 295–312. doi : 10.1146/annurev-immunol-020711-075013 . PMID 22224773.
^ Ho, Pensilvania; Alonso, TA; Gerbing, RB; Pollard, J; Stirewalt, DL; Hurwitz, C; Heerema, NA; Hirsch, B; Raimondi, Carolina del Sur; Lange, B; Franklin, JL; Radich, JP; Meshinchi, S (25 de junio de 2009). "Prevalencia e implicaciones pronósticas de las mutaciones de CEBPA en la leucemia mieloide aguda (LMA) pediátrica: un informe del Grupo de Oncología Infantil". Sangre . 113 (26): 6558–66. doi :10.1182/sangre-2008-10-184747. PMC 2943755 . PMID 19304957.
^ Woolthuis, Carolien M.; Park, Christopher Y. (10 de marzo de 2016). "Compromiso de las células madre / progenitoras hematopoyéticas con el linaje de megacariocitos". Sangre . 127 (10): 1242-1248. doi : 10.1182/sangre-2015-07-607945 . ISSN 0006-4971. PMC 5003506 . PMID 26787736. S2CID 206939258.
^ Thompson, Elizabeth C.; Cobb, Bradley S.; Sabbattini, Pierangela; Meixlsperger, Sonja; Parelho, Vania; Liberg, David; Taylor, Benjamín; Dillon, Niall; Georgopoulos, Katia (1 de marzo de 2007). "Proteínas de unión al ADN de Ikaros como componentes integrales de los circuitos reguladores específicos de la etapa de desarrollo de las células B". Inmunidad . 26 (3): 335–344. doi : 10.1016/j.immuni.2007.02.010 . ISSN 1074-7613. PMID 17363301.
^ Suzuki, Junpei; Maruyama, Saho; Tamauchi, Hidekazu; Kuwahara, Makoto; Horiuchi, Mika; Mizuki, Masumi; Ochi, Mizuki; Sawasaki, Tatsuya; Zhu, Jinfang (1 de abril de 2016). "Gfi1, un represor transcripcional, inhibe la inducción del programa T auxiliar tipo 1 en células T CD4 activadas". Inmunología . 147 (4): 476–487. doi : 10.1111/imm.12580. ISSN 1365-2567. PMC 4799889 . PMID 26749286.
^ Sasaki, Haruka; Kurotaki, Daisuke; Tamura, Tomohiko (1 de abril de 2016). "Regulación del desarrollo de basófilos y mastocitos por factores de transcripción". Alergología Internacional . 65 (2): 127-134. doi : 10.1016/j.alit.2016.01.006 . ISSN 1440-1592. PMID 26972050.
^ O'Brien, P; Morín, P Jr; Ouellette, RJ; Robichaud, GA (15 de diciembre de 2011). "El gen Pax-5: un regulador pluripotente de la diferenciación de células B y la enfermedad cancerosa". Investigación sobre el cáncer . 71 (24): 7345–50. doi : 10.1158/0008-5472.CAN-11-1874 . PMID 22127921.
^ Cobaleda, C; Jochum, W; Busslinger, M (27 de septiembre de 2007). "Conversión de células B maduras en células T mediante desdiferenciación a progenitores no comprometidos" . Naturaleza . 449 (7161): 473–7. Código Bib :2007Natur.449..473C. doi : 10.1038/naturaleza06159. PMID 17851532. S2CID 4414856.
^ Wang, JH; Nichogiannopoulou, A; Wu, L; Sol, L; Sharpe, AH; Bigby, M; Georgopoulos, K (diciembre de 1996). "Defectos selectivos en el desarrollo del sistema linfoide fetal y adulto en ratones con mutación nula de Ikaros". Inmunidad . 5 (6): 537–49. doi : 10.1016/s1074-7613(00)80269-1 . PMID 8986714.
^ Sol, L; Liu, A; Georgopoulos, K (1 de octubre de 1996). "Las interacciones de proteínas mediadas por dedos de zinc modulan la actividad de Ikaros, un control molecular del desarrollo de linfocitos". La Revista EMBO . 15 (19): 5358–69. doi :10.1002/j.1460-2075.1996.tb00920.x. PMC 452279 . PMID 8895580.
^ Schjerven, H; McLaughlin, J; Arenzana, TL; Frietze, S; Cheng, D; Wadsworth, SE; Lawson, GW; Bensinger, SJ; Farnham, PJ; Witte, ON; Smale, ST (octubre de 2013). "Regulación selectiva de la linfopoyesis y leucemogénesis por dedos de zinc individuales de Ikaros". Inmunología de la naturaleza . 14 (10): 1073–83. doi :10.1038/ni.2707. PMC 3800053 . PMID 24013668.
^ Zon, LI (15 de octubre de 1995). "Biología del desarrollo de la hematopoyesis". Sangre (Revisar). 86 (8): 2876–91. doi : 10.1182/sangre.V86.8.2876.2876 . PMID 7579378.
^ Viejo JM (2016). Hematopoyesis en marsupiales. Inmunología comparada y del desarrollo. 58, 40-46. DOI: 10.1016/j.dci.2015.11.009
^ Viejo JM, Deane EM (2000). Desarrollo del sistema inmunológico y protección inmunológica en crías de bolsa de marsupial. Inmunología comparada y del desarrollo. 24(5), 445-454. DOI: 10.1016/S0145-305X(00)00008-2
^ Viejo JM, Deane EM (2003). Los tejidos linfoides e inmunohematopoyéticos de la zarigüeya embrionaria ( Trichosurus vulpecula ). Anatomía y Embriología (ahora llamada Estructura y Función Cerebral). 206(3), 193-197. DOI: 10.1007/s00429-002-0285-2
^ Viejo JM, Selwood L , Deane EM (2003). Una investigación histológica de los tejidos linfoides e inmunohematopoyéticos del dunnart adulto de cara rayada ( Sminthopsis macroura ). Células Tejidos Órganos. 173(2), 115-121. DOI: 10.1159/000068946
Otras lecturas
Godín, Isabelle; Cumano, Ana, eds. (2006). Desarrollo de células madre hematopoyéticas. Saltador. ISBN 978-0-306-47872-7.
enlaces externos
Scholia tiene un perfil de tema para Hematopoyesis .

_diagram_en.svg/1280px-Hematopoiesis_(human)_diagram_en.svg.png)
