Funcionamiento de la GTPasa en el transporte nuclear
Ran ( proteína nuclear relacionada con RA ), también conocida como proteína nuclear de unión a GTP, es una proteína que en los seres humanos está codificada por el gen RAN . Ran es una proteína pequeña de 25 kDa que participa en el transporte hacia dentro y hacia fuera del núcleo celular durante la interfase y también participa en la mitosis . Es un miembro de la superfamilia Ras . [5] [6] [7]
Ran es una proteína G pequeña que es esencial para la translocación de ARN y proteínas a través del complejo de poro nuclear . La proteína Ran también ha estado implicada en el control de la síntesis de ADN y la progresión del ciclo celular, ya que se ha descubierto que las mutaciones en Ran alteran la síntesis de ADN. [8]
Función
Ciclo de ejecución
Representación esquemática del ciclo de Ran
Ran existe en la célula en dos formas unidas a nucleótidos: unida a GDP y unida a GTP . RanGDP se convierte en RanGTP mediante la acción de RCC1 , el factor de intercambio de nucleótidos para Ran. RCC1 también se conoce como RanGEF (factor de intercambio de nucleótidos de guanina Ran). La actividad GTPasa intrínseca de Ran se activa mediante la interacción con la proteína activadora de GTPasa Ran (RanGAP), facilitada por la formación de complejos con la proteína de unión a Ran (RanBP). La activación de GTPasa conduce a la conversión de RanGTP a RanGDP, cerrando así el ciclo de Ran.
Ran puede difundirse libremente dentro de la célula, pero debido a que RCC1 y RanGAP se encuentran en diferentes lugares de la célula, la concentración de RanGTP y RanGDP también difiere localmente, creando gradientes de concentración que actúan como señales para otros procesos celulares. RCC1 está unido a la cromatina y, por lo tanto, se encuentra dentro del núcleo . RanGAP es citoplasmático en levaduras y está unido a la envoltura nuclear en plantas y animales. En células de mamíferos, está modificado por SUMO y se une al lado citoplasmático del complejo del poro nuclear a través de la interacción con la nucleoporina RANBP2 (Nup358). Esta diferencia en la ubicación de las proteínas accesorias en el ciclo de Ran conduce a una alta relación RanGTP a RanGDP dentro del núcleo y una relación RanGTP a RanGDP inversamente baja fuera del núcleo. Además de un gradiente del estado de unión de nucleótidos de Ran, existe un gradiente de la proteína en sí, con una mayor concentración de Ran en el núcleo que en el citoplasma. El RanGDP citoplasmático es importado al núcleo por la pequeña proteína NUTF2 (factor de transporte nuclear 2), donde RCC1 puede luego catalizar el intercambio de GDP por GTP en Ran.
Papel en el transporte nuclear durante la interfase
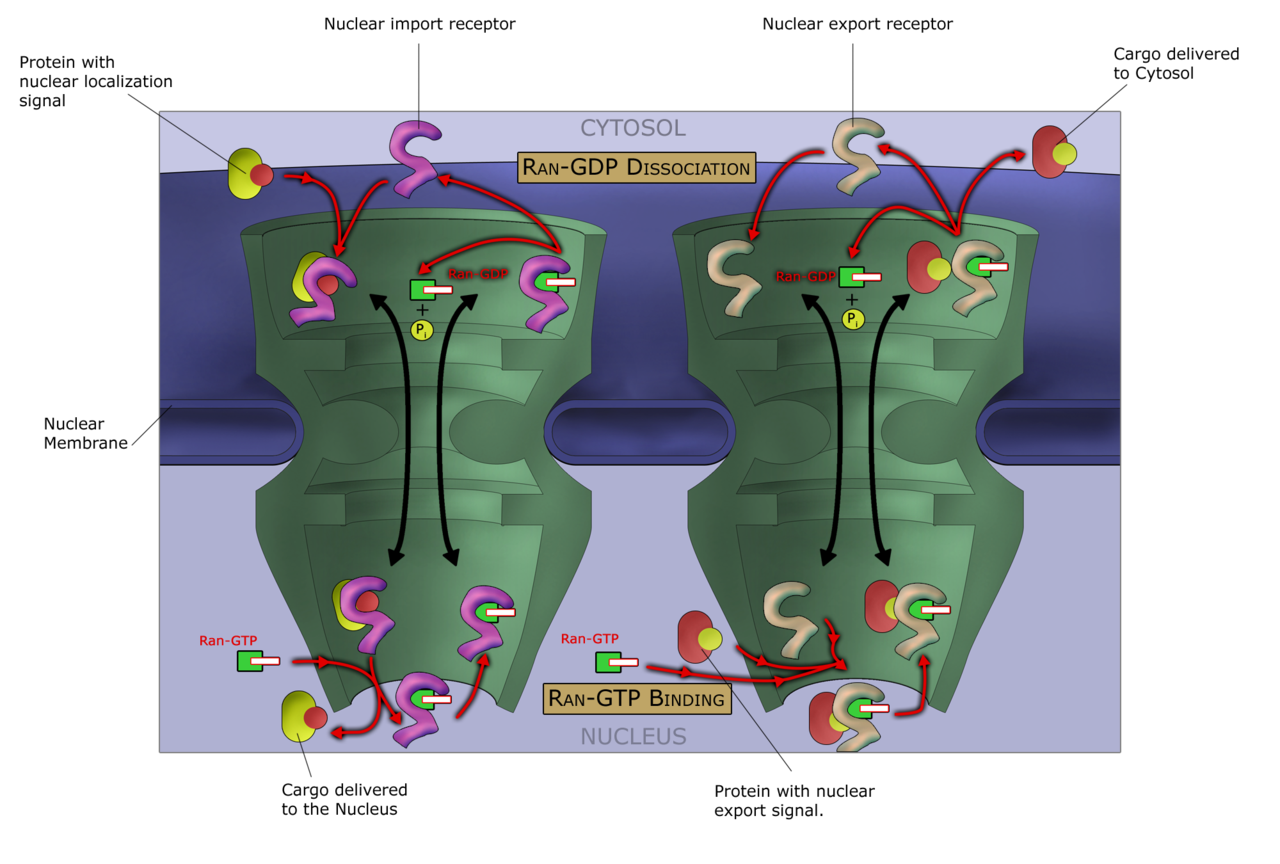
Participación del ciclo Ran en el transporte nucleocitoplasmático en el poro nuclear
Ran participa en el transporte de proteínas a través de la envoltura nuclear al interactuar con las carioferinas y cambiar su capacidad para unirse o liberar moléculas de carga. Las proteínas de carga que contienen una señal de localización nuclear (NLS) son unidas por las importinas y transportadas al núcleo. Dentro del núcleo, RanGTP se une a la importina y libera la carga de importación. La carga que necesita salir del núcleo hacia el citoplasma se une a la exportina en un complejo ternario con RanGTP. Tras la hidrólisis de RanGTP a RanGDP fuera del núcleo, el complejo se disocia y se libera la carga de exportación.
Papel en la mitosis
Durante la mitosis, el ciclo Ran está involucrado en el ensamblaje del huso mitótico y el reensamblaje de la envoltura nuclear después de que los cromosomas se han separado. [9] [10] Durante la profase , el gradiente pronunciado en la relación RanGTP-RanGDP en los poros nucleares se rompe a medida que la envoltura nuclear se vuelve permeable y se desmonta. La concentración de RanGTP se mantiene alta alrededor de los cromosomas ya que RCC1, un factor de intercambio de nucleótidos, permanece unido a la cromatina . [11] RanBP2 (Nup358) y RanGAP se mueven a los cinetocoros donde facilitan la unión de las fibras del huso a los cromosomas. Además, RanGTP promueve el ensamblaje del huso por mecanismos similares a los mecanismos de transporte nuclear: la actividad de los factores de ensamblaje del huso como NuMA y TPX2 es inhibida por la unión a las importinas. Al liberar importinas, RanGTP activa estos factores y, por lo tanto, promueve el ensamblaje del huso mitótico . En la telofase , la hidrólisis de RanGTP y el intercambio de nucleótidos son necesarios para la fusión de vesículas en las envolturas nucleares reformadoras de los núcleos hijos.
Ran y el receptor de andrógenos
El RAN es un coactivador del receptor de andrógenos (ARA24) que se une de forma diferencial con diferentes longitudes de poliglutamina dentro del receptor de andrógenos. La expansión de repeticiones de poliglutamina en el AR está vinculada a la atrofia muscular espinal y bulbar (enfermedad de Kennedy). La coactivación del AR por el RAN disminuye con la expansión de poliglutamina dentro del AR, y esta débil coactivación puede conducir a una insensibilidad parcial a los andrógenos durante el desarrollo de la atrofia muscular espinal y bulbar. [12] [13]
^ abc GRCh38: Lanzamiento de Ensembl 89: ENSG00000132341 – Ensembl , mayo de 2017
^ abc GRCm38: Lanzamiento de Ensembl 89: ENSMUSG00000029430 – Ensembl , mayo de 2017
^ "Referencia de PubMed humana:". Centro Nacional de Información Biotecnológica, Biblioteca Nacional de Medicina de EE. UU .
^ "Referencia PubMed de ratón:". Centro Nacional de Información Biotecnológica, Biblioteca Nacional de Medicina de EE. UU . .
^ Moore MS, Blobel G (mayo de 1994). "Proteína AG implicada en el transporte nucleocitoplasmático: el papel de Ran". Trends Biochem. Sci . 19 (5): 211–6. doi :10.1016/0968-0004(94)90024-8. PMID 7519373.
^ Avis JM, Clarke PR (octubre de 1996). "Ran, una GTPasa implicada en procesos nucleares: sus reguladores y efectores". J. Cell Sci . 109 (10): 2423–7. doi :10.1242/jcs.109.10.2423. PMID 8923203.
^ Dasso M, Pu RT (agosto de 1998). "Transporte nuclear: ¿gestionado por Ran?". Am. J. Hum. Genet . 63 (2): 311–6. doi :10.1086/301990. PMC 1377330. PMID 9683621 .
^ Sazer S, Dasso M (abril de 2000). "El decatlón de Ran: múltiples roles de Ran". J. Cell Sci . 113 (7): 1111–8. doi :10.1242/jcs.113.7.1111. PMID 10704362.
^ Gruss OJ, Vernos I (septiembre de 2004). "El mecanismo de ensamblaje del huso: funciones de Ran y su diana TPX2". J. Cell Biol . 166 (7): 949–55. doi :10.1083/jcb.200312112. PMC 2172015. PMID 15452138 .
^ Ciciarello M, Mangiacasale R, Lavia P (agosto de 2007). "Control espacial de la mitosis por la GTPasa Ran". Cell. Mol. Life Sci . 64 (15): 1891–914. doi :10.1007/s00018-007-6568-2. PMC 11136129. PMID 17483873. S2CID 8687055 .
^ Carazo-Salas RE, Guarguaglini G, Gruss OJ, Segref A, Karsenti E, Mattaj IW (julio de 1999). "La generación de Ran unida a GTP por RCC1 es necesaria para la formación del huso mitótico inducida por cromatina". Nature . 400 (6740): 178–81. Bibcode :1999Natur.400..178C. doi :10.1038/22133. PMID 10408446. S2CID 4417176.
^ Hsiao PW, Lin DL, Nakao R, Chang C (julio de 1999). "La relación entre la enfermedad neuronal de Kennedy y ARA24, el primer coactivador asociado a la región de poliglutamina del receptor de andrógenos identificado". J. Biol. Chem . 274 (29): 20229–34. doi : 10.1074/jbc.274.29.20229 . PMID 10400640.
^ Sampson ER, Yeh SY, Chang HC, Tsai MY, Wang X, Ting HJ, Chang C (2001). "Identificación y caracterización de correguladores asociados al receptor de andrógenos en células de cáncer de próstata". J. Biol. Regul. Homeost. Agents . 15 (2): 123–9. PMID 11501969.
^ abc Plafker K, Macara IG (2000). "Transporte nucleocitoplasmático facilitado de la proteína de unión a Ran RanBP1". Mol. Cell. Biol . 20 (10): 3510–21. doi :10.1128/MCB.20.10.3510-3521.2000. PMC 85643. PMID 10779340 .
^ Kutay U, Izaurralde E, Bischoff FR, Mattaj IW, Görlich D (1997). "Los mutantes dominantes negativos de la importina beta bloquean múltiples vías de importación y exportación a través del complejo del poro nuclear". EMBO J . 16 (6): 1153–63. doi :10.1093/emboj/16.6.1153. PMC 1169714 . PMID 9135132.
^ Percipalle P, Clarkson WD, Kent HM, Rhodes D, Stewart M (1997). "Interacciones moleculares entre el heterodímero de importina alfa/beta y las proteínas implicadas en la importación de proteínas nucleares de vertebrados". J. Mol. Biol . 266 (4): 722–32. doi :10.1006/jmbi.1996.0801. PMID 9102465.
^ Roig J, Mikhailov A, Belham C, Avruch J (2002). "Nercc1, una quinasa de la familia NIMA de mamíferos, se une a la GTPasa Ran y regula la progresión mitótica". Genes Dev . 16 (13): 1640–58. doi :10.1101/gad.972202. PMC 186374 . PMID 12101123.
^ Cushman I, Bowman BR, Sowa ME, Lichtarge O, Quiocho FA, Moore MS (2004). "Identificación computacional y bioquímica de un sitio de unión de complejo de poro nuclear en el transportador nuclear NTF2". J. Mol. Biol . 344 (2): 303–10. doi :10.1016/j.jmb.2004.09.043. PMID 15522285.
^ Stewart M, Kent HM, McCoy AJ (1998). "Base estructural para el reconocimiento molecular entre el factor de transporte nuclear 2 (NTF2) y la forma unida a GDP de la GTPasa Ran de la familia Ras". J. Mol. Biol . 277 (3): 635–46. doi :10.1006/jmbi.1997.1602. PMID 9533885.
^ ab Steggerda SM, Paschal BM (2000). "La proteína Mog1 de mamíferos es un factor de liberación de nucleótidos de guanina para Ran". J. Biol. Chem . 275 (30): 23175–80. doi : 10.1074/jbc.C000252200 . PMID 10811801.
^ ab Ren M, Villamarin A, Shih A, Coutavas E, Moore MS, LoCurcio M, Clarke V, Oppenheim JD, D'Eustachio P, Rush MG (1995). "Dominios separados de la GTPasa Ran interactúan con diferentes factores para regular la importación de proteínas nucleares y el procesamiento del ARN". Mol. Cell. Biol . 15 (4): 2117–24. doi :10.1128/MCB.15.4.2117. PMC 230439. PMID 7891706 .
^ Hillig RC, Renault L, Vetter IR, Drell T, Wittinghofer A, Becker J (1999). "La estructura cristalina de rna1p: un nuevo pliegue para una proteína activadora de GTPasa". Mol. Cell . 3 (6): 781–91. doi : 10.1016/S1097-2765(01)80010-1 . PMID 10394366.
^ Becker J, Melchior F, Gerke V, Bischoff FR, Ponstingl H, Wittinghofer A (1995). "RNA1 codifica una proteína activadora de GTPasa específica para Gsp1p, el homólogo Ran/TC4 de Saccharomyces cerevisiae". J. Biol. Chem . 270 (20): 11860–5. doi : 10.1074/jbc.270.20.11860 . PMID 7744835.
^ Bischoff FR, Klebe C, Kretschmer J, Wittinghofer A, Ponstingl H (1994). "RanGAP1 induce la actividad GTPasa de Ran relacionada con Ras nuclear". Proc. Natl. Sci. EE. UU . . 91 (7): 2587–91. Bibcode :1994PNAS...91.2587B. doi : 10.1073/pnas.91.7.2587 . PMC 43414 . PMID 8146159.
^ Renault L, Kuhlmann J, Henkel A, Wittinghofer A (2001). "Base estructural para el intercambio de nucleótidos de guanina en Ran por el regulador de la condensación cromosómica (RCC1)". Cell . 105 (2): 245–55. doi : 10.1016/S0092-8674(01)00315-4 . PMID 11336674. S2CID 12827419.
^ Azuma Y, Renault L, García-Ranea JA, Valencia A, Nishimoto T, Wittinghofer A (1999). "Modelo de la interacción ran-RCC1 usando experimentos bioquímicos y de acoplamiento". J. Mol. Biol . 289 (4): 1119–30. doi :10.1006/jmbi.1999.2820. PMID 10369786.
^ Chook YM, Blobel G (1999). "Estructura del complejo de transporte nuclear carioferina-beta2-Ran x GppNHp". Nature . 399 (6733): 230–7. Código Bibliográfico :1999Natur.399..230C. doi :10.1038/20375. PMID 10353245. S2CID 4413233.
^ ab Shamsher MK, Ploski J, Radu A (2002). "La carioferina beta 2B participa en la exportación de ARNm desde el núcleo". Proc. Natl. Sci. EE. UU . . 99 (22): 14195–9. Bibcode :2002PNAS...9914195S. doi : 10.1073/pnas.212518199 . PMC 137860 . PMID 12384575.
^ Tickenbrock L, Cramer J, Vetter IR, Muller O (2002). "La región de la espiral enrollada (aminoácidos 129-250) de la proteína supresora de tumores adenomatosa poliposis coli (APC). Su estructura y su interacción con la región de mantenimiento de cromosomas 1 (Crm-1)". J. Biol. Chem . 277 (35): 32332–8. doi : 10.1074/jbc.M203990200 . PMID 12070164.
^ Fornerod M, Ohno M, Yoshida M, Mattaj IW (1997). "CRM1 es un receptor de exportación para señales de exportación nuclear ricas en leucina". Cell . 90 (6): 1051–60. doi : 10.1016/S0092-8674(00)80371-2 . PMID 9323133. S2CID 15119502.
^ Brownawell AM, Macara IG (2002). "Exportin-5, una nueva carioferina, media la exportación nuclear de proteínas de unión a ARN de doble cadena". J. Cell Biol . 156 (1): 53–64. doi :10.1083/jcb.200110082. PMC 2173575. PMID 11777942 .
^ Ørom UA, Nielsen FC, Lund AH (2008). "El microARN-10a se une al 5'UTR de los ARNm de las proteínas ribosómicas y mejora su traducción". Mol Cell . 30 (4): 460–71. doi : 10.1016/j.molcel.2008.05.001 . PMID 18498749.
Patel SS. "Animaciones de factores de transporte nuclear, incluido Ran (1 de 2 páginas)". Animaciones de transporte nuclear . Archivado desde el original el 2009-02-07 . Consultado el 2008-06-12 .
Patel SS. "Animaciones de factores de transporte nuclear, incluido Ran (2 de 2 páginas)". Animaciones de transporte nuclear . Archivado desde el original el 2009-02-07 . Consultado el 2008-06-12 .

{kind=link}