Una célula hipercompleja (actualmente llamada célula de terminación terminal ) es un tipo de neurona de procesamiento visual en la corteza cerebral de los mamíferos . Descubiertas inicialmente por David Hubel y Torsten Wiesel en 1965, las células hipercomplejas se definen por la propiedad de terminación terminal, que es una disminución en la fuerza de disparo con estímulos cada vez más grandes . La sensibilidad a la longitud del estímulo va acompañada de selectividad para la orientación, el movimiento y la dirección específicos de los estímulos. Por ejemplo, una célula hipercompleja solo puede responder a una línea a 45˚ que viaja hacia arriba. Alargar la línea daría como resultado una respuesta proporcionalmente más débil. En última instancia, las células hipercomplejas pueden proporcionar un medio para que el cerebro perciba visualmente las esquinas y las curvas del entorno al identificar los extremos de un estímulo determinado. [1]
Las células hipercomplejas se caracterizaron originalmente como la clase superordinada de células de procesamiento visual por encima de las células complejas y simples . Mientras que las células complejas eran sensibles a estímulos en movimiento de orientaciones específicas que viajan en una dirección específica, las células simples solo respondían a estímulos lineales orientados correctamente. No se creía que las células simples ni las complejas mostraran detención final. Asimismo, se creía que la detención final estaba restringida a áreas visuales de orden superior ( área 18 de Brodmann y área 19 de Brodmann ), pero más tarde se descubrió que también existía en la corteza visual primaria ( área 17 de Brodmann ). En 1968, Geoffrey Henry y Bogdan Dreher descubrieron células simples y complejas con propiedades de detención final. Posteriormente, las células hipercomplejas ya no se reconocieron como una clase distinta, sino más bien como un subtipo de células simples y complejas. Actualmente, las células simples con detención final y las células complejas con detención final son los términos de elección para describir las neuronas con propiedades de detención final. [1]


En la década de 1950, el conocimiento de la función cortical era relativamente limitado. Sin embargo, hacia finales de la década, se estaba estableciendo la plataforma para comprender la corteza. Las investigaciones sobre la localización de la función, así como la aparición de registros unicelulares de neuronas, fomentaron una mayor comprensión del procesamiento de la información desde la sensación hasta la percepción. Con referencia a la visión, Stephen Kuffler descubrió áreas de la retina , denominadas campos receptivos , que, al ser estimuladas, influirían en la activación de las células ganglionares . [2] Estos campos comprendían dos capas concéntricas, una excitatoria y otra inhibidora. Un tipo de campo receptivo se describió como centrado, que contiene un centro excitatorio y un entorno inhibidor, mientras que el otro tipo se denominó descentrado, que contiene un centro inhibidor y un entorno excitatorio. Se descubrieron campos receptivos similares en el núcleo geniculado lateral (LGN). [2]
Dos estudiantes de doctorado del laboratorio de Kuffler en la Universidad Johns Hopkins , David Hubel y Torsten Wiesel, recibieron el encargo de ampliar su trabajo desde las células ganglionares de la retina hasta la corteza visual . Hubel y Wiesel comenzaron a registrar células de la corteza mientras presentaban puntos de luz como estímulos. Al principio, los dos no habían logrado producir ninguna grabación prometedora, ya que las células no respondían a los estímulos dados. Sin embargo, al insertar la lámina de vidrio en el proyector, se obtuvo inmediatamente una señal fuerte. Casualmente, Hubel y Wiesel descubrieron que la célula no respondía a los puntos, sino a los bordes, es decir, a la sombra de la lámina cuando se colocaba en el proyector. [2] [3]
Hubel y Wiesel llamarían más tarde a esta célula una célula compleja , incorporándola a una jerarquía de células de procesamiento visual descubiertas posteriormente, que incluían las células centro-envolventes, simples, complejas e hipercomplejas (distinguibles por los campos receptivos) [4].
Tras su hallazgo inicial, Hubel y Wiesel descubrieron la presencia de una variedad de células de procesamiento visual, cada una con propiedades de campo receptivo únicas. En el nivel más bajo y más simple de la jerarquía se encuentran las células centro-envolventes mencionadas anteriormente del ganglio retiniano y el LGN. A continuación, dentro de la corteza visual, se encuentran las células simples. [4] Las células simples existen dentro de la corteza visual primaria (Área 17 de Brodmann). Estas células se encuentran específicamente en la capa IV , en la que terminan la mayoría de las proyecciones salientes del LGN. [4] Los campos receptivos de las células simples son no concéntricos y lineales, en los que existen regiones excitatorias e inhibidoras adyacentes entre sí. Por lo tanto, se produce una respuesta por estímulos lineales estacionarios. Además, las regiones exhiben cancelación mutua (antagonismo) y producen respuestas más fuertes a medida que los estímulos llenan más espacio ( suma espacial ). Una característica distintiva de las células simples es que sus respuestas muestran orientación y selectividad posicional. Esto significa que una célula simple se activa en una orientación óptima. Las respuestas obtenidas se debilitan progresivamente a medida que la orientación del estímulo cambia de forma subóptima y dejan de activarse cuando se encuentra a 90° de la orientación óptima. La selectividad posicional simplemente se refiere a la receptividad de la célula a la posición del estímulo dentro de una parte o de la totalidad de las regiones excitatorias/inhibitorias. En consecuencia, existen campos receptivos celulares simples en una variedad de geometrías y tamaños diferentes para todas las orientaciones y posiciones posibles en el campo visual. Se presume que múltiples campos receptivos concéntricos del LGN convergen en una línea para desarrollar un único campo receptivo simple. [4] [5]
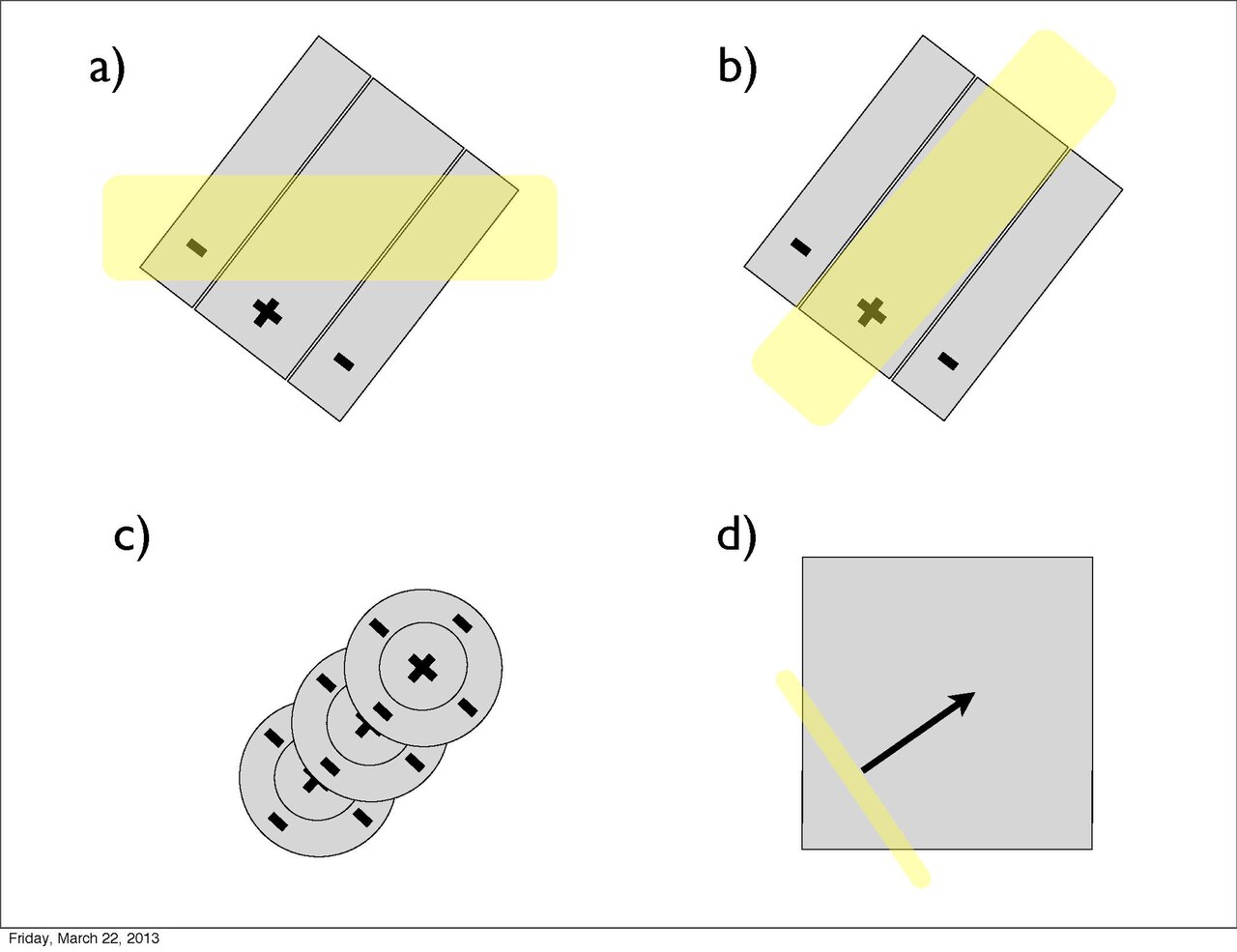
Además de las células simples, existen células complejas, que son el tipo más común en la corteza visual primaria (pero también se encuentran en el área 18 de Brodmann). Al igual que las células simples, los campos receptivos de las células complejas son selectivos en cuanto a la orientación. Sin embargo, a diferencia de las células simples, las células complejas no responden a estímulos estacionarios. Para producir una respuesta sostenida, el estímulo debe moverse a través del campo receptivo. La selectividad de movimiento de las células complejas significa que se produce una respuesta en una amplia gama de posiciones de estímulo. Una cantidad sustancial de células complejas también muestran selectividad direccional, de modo que el movimiento en una sola dirección produce una respuesta óptima. La arquitectura cortical de las células complejas consiste en células simples adyacentes convergentes con campos receptivos que muestran la misma selectividad de orientación. Para explicar la selectividad de movimiento de las células complejas, Hubel y Wiesel postularon que el sistema de células simples solo produce una respuesta breve a estímulos estacionarios (es decir, la respuesta se adapta ). En consecuencia, se requieren estimulaciones sucesivas que avancen a través del campo receptivo complejo para producir una respuesta sostenida; por lo tanto, se produce selectividad de movimiento. [4]
Aunque las definiciones anteriores, establecidas por Hubel y Wiesel, son las más aceptadas, algunos de sus contemporáneos habían distinguido inicialmente las clases según diferentes criterios. En resumen, Hubel y Wiesel identificaron las células simples mediante regiones excitatorias e inhibidoras claramente separadas que respondían a estímulos estacionarios. En contraste, Peter Bishop utilizó otros criterios e incluyó los estímulos en movimiento dentro de la definición de células simples. [1]
Además de los esquemas de cableado de Hubel y Wiesel, se han propuesto múltiples arquitecturas alternativas y complementarias para explicar los campos receptivos de células simples y complejas:
En 1965, se encontró el siguiente tipo de célula en la jerarquía de procesamiento visual de Hubel y Wiesel, la célula hipercompleja, dentro de las áreas 18 y 19 de Brodmann. Tras su descubrimiento, las células hipercomplejas se definieron como "todas las células que superan a las células complejas en complejidad de comportamiento". [7] Las células hipercomplejas mostraron una selectividad similar a la de las células complejas, respondiendo al movimiento de un estímulo de una orientación específica en una dirección específica.
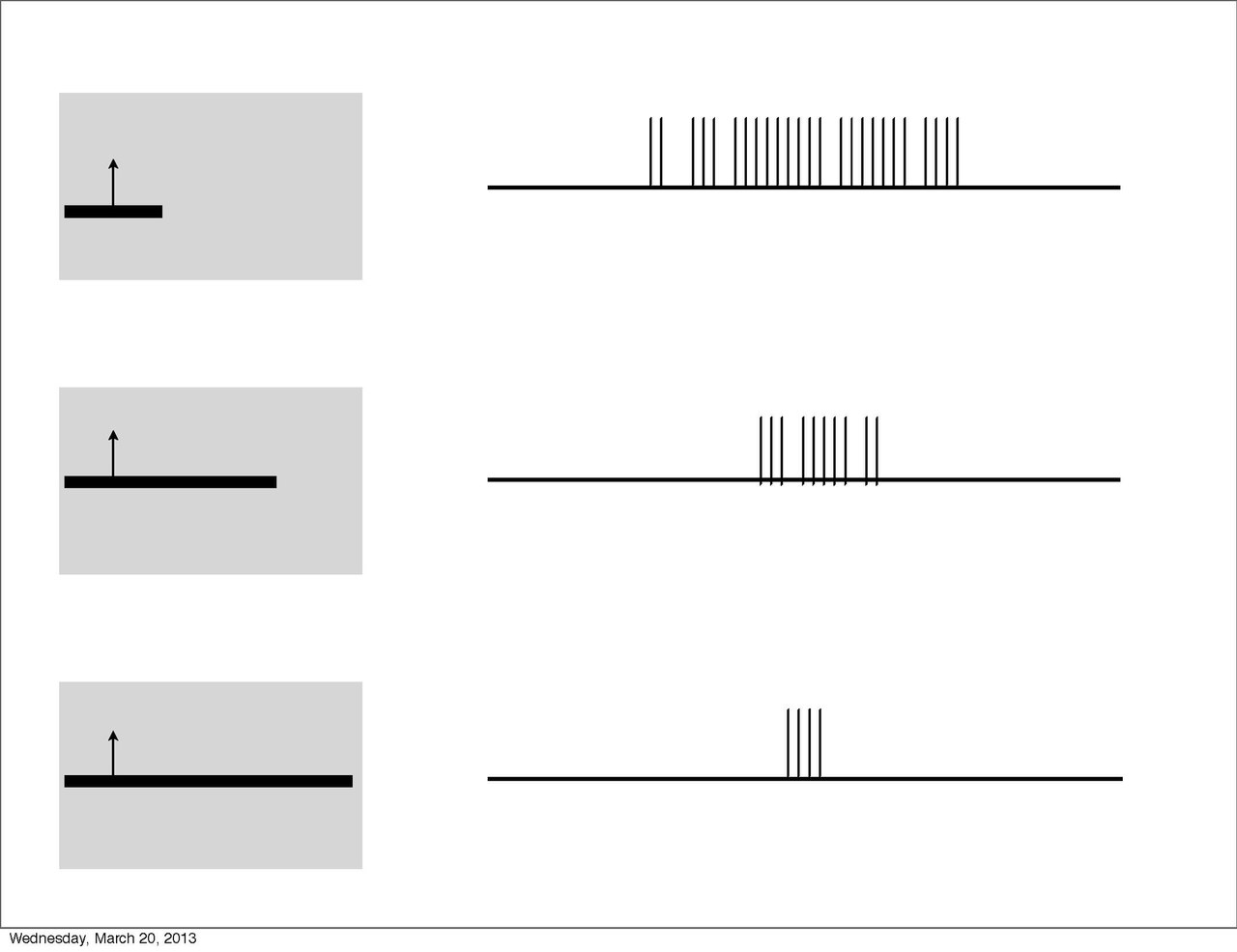
Además, al igual que las células de procesamiento subordinadas, el aumento de la iluminación en una región particular provocó respuestas más fuertes (es decir, suma espacial). Sin embargo, esta suma se limitaba a estímulos de un tamaño limitado. Al extenderse más allá de una longitud específica, la respuesta se volvería progresivamente más débil. Este fenómeno se denomina detención final y es la propiedad definitoria de las células hipercomplejas. Hubel y Wiesel caracterizan estos campos receptivos como que contienen regiones activadoras y antagonistas (similares a las regiones excitatorias/inhibitorias). Por ejemplo, la mitad izquierda de un campo receptivo puede ser la región activadora, mientras que la región antagonista se encuentra a la derecha. En consecuencia, la célula hipercompleja responderá, con suma espacial, a los estímulos en el lado izquierdo (dentro de la región activadora) en la medida en que no se extienda más hacia el lado derecho (región antagonista). Este campo receptivo se describiría como detenido en un extremo (es decir, el derecho). De manera similar, los campos receptivos hipercomplejos pueden detenerse en ambos extremos. En este caso, un estímulo que se extiende demasiado en cualquier dirección (por ejemplo, demasiado a la izquierda o demasiado a la derecha) comenzará a estimular la región antagonista y reducirá la fuerza de la señal de la célula. [7] Nótese que las células hipercomplejas también son selectivas a la orientación, el movimiento y la dirección. De hecho, la región activadora tendrá la misma selectividad de orientación que la región antagonista. Por lo tanto, solo una línea que se extienda hacia la región antagonista disminuirá la fuerza de la respuesta, en lugar de otra línea orientada de manera diferente. Un posible esquema para el cableado de células hipercomplejas podría comprender la entrada excitatoria de una célula compleja dentro de la región activadora y la entrada inhibidora de células complejas en las regiones antagonistas periféricas. [4] [8]
Poco después de que Hubel y Wiesel incluyeran la hipercomplejidad en su versión de la jerarquía de procesamiento visual, se planteó la noción de una clase de células hipercomplejas. En 1968, Geoffrey Henry y Bogdan Dreher descubrieron células simples y complejas en el área 17 de Brodmann que exhibían propiedades de detención en los extremos. [9] En lugar de caracterizar la detención en los extremos como algo exclusivo de una clase supraordinada de neuronas, era más apropiado atribuirla como una propiedad de las células simples y complejas. [2] Sólo unos pocos años después, Charles Gilbert, un estudiante de posgrado de Hubel y Wiesel, había confirmado la detención en los extremos en la corteza visual primaria. [10] En consecuencia, se introdujeron los términos de célula simple de detención en los extremos y de célula compleja de detención en los extremos en lugar de célula hipercompleja. Las células hipercomplejas descritas por Hubel y Wiesel anteriormente eran probablemente un conjunto de células complejas de detención en los extremos. [11] En su discurso de entrega del Premio Nobel , Hubel explicó que la jerarquía de las células de procesamiento visual resultó ser más complicada y amorfa de lo que se creía inicialmente, y señaló que el tema comenzaba a parecerse a una “jungla”. [2]

En última instancia, estas células contribuyen a los mecanismos que subyacen a la percepción visual. Una célula simple con un extremo finalizado mostrará selectividad en cuanto a longitud y orientación. En términos de arquitectura cortical, puede recibir información de células simples ordinarias de orientación idéntica. [4] Por ejemplo, la región activadora podría consistir en una célula simple que envía información excitatoria, mientras que la región antagonista podría consistir en células simples que proporcionan información inhibidora. Una célula compleja con un extremo finalizado seleccionaría orientación, movimiento y dirección, pero también longitud. Podría recibir información de un conjunto de células complejas, de manera similar al esquema mencionado anteriormente. La región activadora podría consistir en una célula compleja que envía información excitatoria y la región antagonista podría consistir en células complejas que envían información inhibidora. [4]
El estímulo óptimo para cualquier célula detenida en un extremo es uno de longitud limitada. Esto se traduce en una capacidad para identificar esquinas (para células detenidas en un extremo) y curvas (para células detenidas en ambos extremos). [4] [12] De la misma manera, la corteza percibe escenas visuales con énfasis en los bordes y límites de los objetos. [13] Las células de procesamiento visual en la corteza responden muy mal a la luz difusa, pero de manera óptima a las líneas. Por ejemplo, una célula simple solo se activará débilmente si está completamente iluminada porque se estimularán tanto las regiones excitatorias como las inhibidoras.
Si el objeto fuera un cuadrado, por ejemplo, las células simples con campos receptivos que correspondieran al interior del cuadrado no serían estimuladas. Sin embargo, una célula simple con un campo receptivo que correspondiera al borde del cuadrado sería estimulada siempre que el borde se encuentre dentro de su región excitatoria. Siguiendo el mismo ejemplo, las células complejas responderían débilmente al interior pero fuertemente a un borde apropiado. Por último, las células con extremos redondeados también serían estimuladas por las esquinas del cuadrado. Una célula con extremos redondeados no respondería a un borde en el lado del cuadrado porque la línea estimularía simultáneamente tanto la región activadora como la antagonista. Por ejemplo, una célula que se detuviera en el extremo derecho (es decir, la región antagonista de la derecha) sería estimulada por la esquina derecha. Aunque percibir un cuadrado implica mucho más que las contribuciones de las células simples y complejas, este ejemplo ilustra que los bordes y los límites de un estímulo (sin entrada desde el interior) son suficientes para interpretar su forma. Por lo tanto, el mecanismo de centrarse en los bordes para traducir la activación en percepción es un uso eficiente de los recursos neuronales.
Aunque las células con terminación terminal son un fenómeno de la corteza visual de los mamíferos, se han descubierto células que exhiben propiedades de terminación terminal en una variedad de otras especies . Por ejemplo, los detectores de movimiento de objetivos pequeños (STMD, por sus siglas en inglés) de muchos insectos seleccionan objetivos pequeños en movimiento, pero se inhiben o no responden a estímulos más grandes. Los STMD se utilizan para discernir insectos en movimiento del desorden circundante y, por lo tanto, son vitales para los comportamientos de persecución. [14]
Además de investigar los efectos integradores de la detención final en la percepción visual, los investigadores están incorporando células de detención final (y otras células de procesamiento visual) en modelos computacionales que simulan la representación jerárquica de la forma en el cerebro. [15] [16]