El complejo de silenciamiento inducido por ARN , o RISC , es un complejo multiproteico , específicamente una ribonucleoproteína , que funciona en el silenciamiento génico a través de una variedad de vías a nivel transcripcional y traduccional. [1] Utilizando fragmentos de ARN monocatenario (ssRNA), como microARN (miRNA), o ARN interferente pequeño bicatenario (siRNA), el complejo funciona como una herramienta clave en la regulación génica. [2] La hebra simple de ARN actúa como una plantilla para que RISC reconozca la transcripción de ARN mensajero complementario (ARNm) . Una vez encontrada, una de las proteínas en RISC, Argonaute , activa y escinde el ARNm. Este proceso se llama interferencia de ARN (RNAi) y se encuentra en muchos eucariotas ; es un proceso clave en la defensa contra infecciones virales , ya que se desencadena por la presencia de ARN bicatenario (dsRNA). [3] [4] [1]
Las células S2 se transfectaron con el ARN de doble cadena de ciclina E de Drosophila . La ciclina E es un gen esencial para la progresión del ciclo celular hacia la fase S. El ARN de doble cadena de ciclina E detuvo el ciclo celular en la fase G1 (antes de la fase S). Por lo tanto, el ARNi puede dirigirse a genes endógenos .
Además, el dsRNA de ciclina E solo disminuyó el ARN de ciclina E; también se demostró un resultado similar utilizando dsRNA correspondiente a la ciclina A que actúa en las fases S, G 2 y M del ciclo celular. Esto muestra el sello característico del RNAi: los niveles reducidos de ARNm corresponden a los niveles de dsRNA agregados.
Para probar si su observación de niveles disminuidos de ARNm era el resultado de que el ARNm fuera atacado directamente (como lo sugerían los datos de otros sistemas), las células S2 de Drosophila se transfectaron con dsRNA de ciclina E de Drosophila o dsRNA de lacZ y luego se incubaron con ARNm sintéticos para ciclina E o lacZ .
Las células transfectadas con dsRNA de ciclina E solo mostraron degradación en las transcripciones de ciclina E; las transcripciones de lacZ fueron estables. Por el contrario, las células transfectadas con dsRNA de lacZ solo mostraron degradación en las transcripciones de lacZ y no en las transcripciones de ciclina E. Sus resultados llevaron a Hannon y sus colegas a sugerir que el RNAi degrada el ARNm objetivo a través de una " actividad nucleasa específica de secuencia ". Llamaron a la enzima nucleasa RISC. [5] Más tarde, Devanand Sarkar y sus colegas Prasanna K. Santhekadur y Byoung Kwon Yoo en la Universidad Commonwealth de Virginia dilucidaron la actividad RISC y su mecanismo molecular en las células cancerosas e identificaron otro nuevo componente del RISC, llamado AEG-1 [47].
Función en la interferencia del ARN
El dominio PIWI de una proteína Argonauta en complejo con ARN bicatenario.
Incorporación de siRNA/miRNA
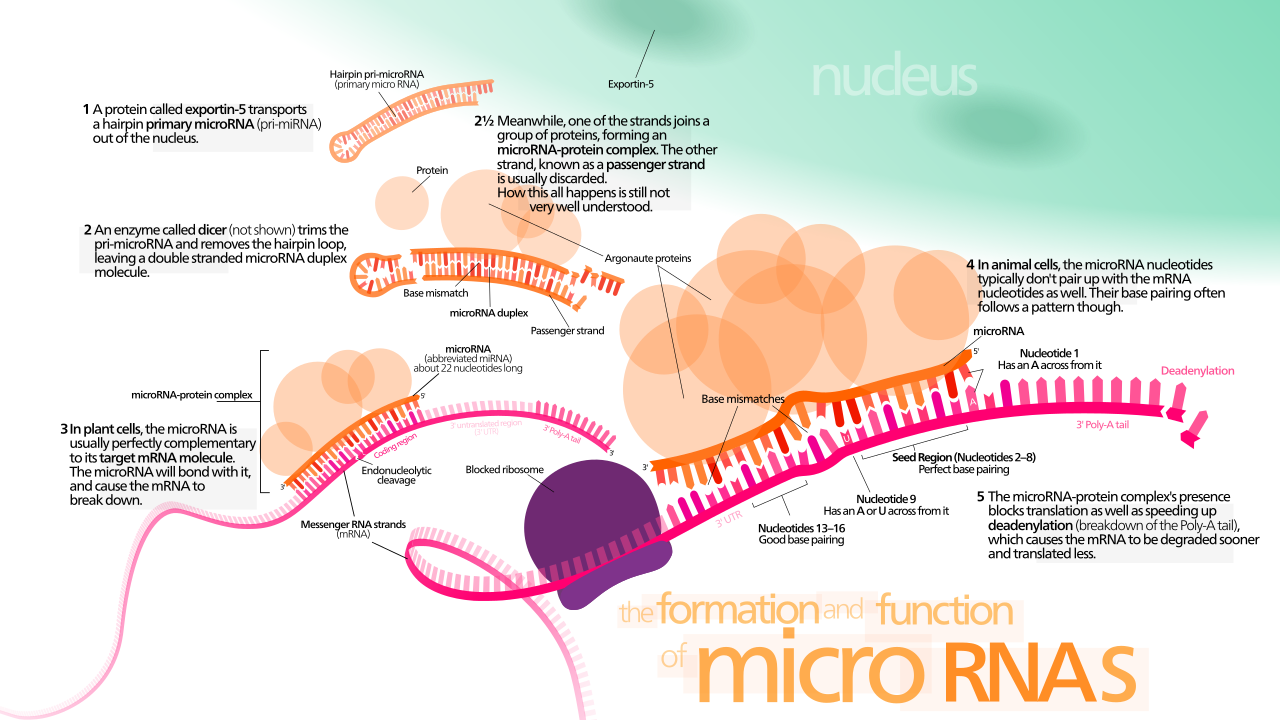
El Dicer de la ARNasa III es un miembro fundamental de RISC que inicia el proceso de interferencia del ARN mediante la producción de ARNi de doble cadena o miARN de cadena sencilla. La escisión enzimática del ARNbc dentro de la célula produce fragmentos cortos de ARNi de 21 a 23 nucleótidos de longitud con un saliente 3' de dos nucleótidos . [6] [7] Dicer también procesa el pre-miARN, que forma una estructura de bucle de horquilla para imitar al ARNbc, de manera similar. Los fragmentos de ARNbc se cargan en RISC y cada cadena tiene un destino diferente según el fenómeno de la regla de asimetría, la selección de una cadena como cadena guía sobre la otra según la estabilidad termodinámica. [8] [9] [10] [11] El miARN o el ARNi recién generados actúan como secuencias guía de cadena sencilla para que RISC dirija el ARNm para su degradación. [12] [13]
La cadena con el extremo 5' menos estable termodinámicamente es seleccionada por la proteína Argonaute e integrada en RISC. [11] [14] Esta cadena se conoce como la cadena guía y se dirige al ARNm para su degradación.
La otra hebra, conocida como hebra pasajera, es degradada por RISC. [15]
Parte de la vía de interferencia de ARN con las diferentes formas en que RISC puede silenciar genes a través de su ARN mensajero.
Regulación genética
AGO2 (gris) en complejo con un microARN (azul claro) y su ARNm diana (azul oscuro)
Las principales proteínas de RISC, Ago2, SND1 y AEG-1, actúan como contribuyentes cruciales a la función de silenciamiento genético del complejo. [16]
La función más conocida de RISC es la degradación del ARNm objetivo, lo que reduce los niveles de transcripción disponibles para ser traducidos por los ribosomas . La escisión endonucleolítica del ARNm complementario a la cadena guía de RISC por la proteína Argonaute es la clave para la iniciación de la interferencia de ARN. [18] Hay dos requisitos principales para que se produzca la degradación del ARNm:
una coincidencia complementaria casi perfecta entre la cadena guía y la secuencia de ARNm objetivo, y
una proteína Argonauta catalíticamente activa, llamada "slicer", para cortar el ARNm objetivo. [1]
Existen dos vías principales de degradación del ARNm una vez que se ha producido la escisión. Ambas se inician mediante la degradación de la cola poli(A) del ARNm , lo que da como resultado la eliminación de la tapa 5' del ARNm.
La degradación de 3' a 5' de la transcripción la lleva a cabo el exosoma y el complejo Ski . [18]
Represión traduccional
El RISC puede modular la carga de ribosoma y factores accesorios en la traducción para reprimir la expresión de la transcripción de ARNm unida. La represión de la traducción solo requiere una coincidencia parcial de la secuencia entre la cadena guía y el ARNm objetivo. [1]
La traducción se puede regular en la etapa inicial mediante:
Se ha demostrado que RITS dirige la formación de heterocromatina en los centrómeros a través del reconocimiento de repeticiones centroméricas. Mediante el apareamiento de bases de ARNi (cadena guía) con secuencias de cromatina objetivo, se pueden reclutar enzimas modificadoras de histonas. [25]
El mecanismo no se entiende bien; sin embargo, los RITS degradan las transcripciones nacientes de ARNm. Se ha sugerido que este mecanismo actúa como un " ciclo de retroalimentación que se refuerza a sí mismo ", ya que las transcripciones nacientes degradadas son utilizadas por la ARN polimerasa dependiente de ARN (RdRp) para generar más ARNi. [26]
En Schizosaccharomyces pombe y Arabidopsis , el procesamiento de los ARN de doble cadena en ARNi por las ARNasas Dicer puede iniciar una vía de silenciamiento génico mediante la formación de heterocromatina. Una proteína Argonaute conocida como AGO4 interactúa con los ARN pequeños que definen secuencias heterocromáticas. Una histona metil transferasa (HMT), H3K9 , metila la histona H3 y recluta proteínas de cromodominio a los sitios de metilación. La metilación del ADN mantiene el silenciamiento de los genes a medida que las secuencias de heterocromatina pueden establecerse o propagarse. [27]
Eliminación de ADN
El ARNi generado por los RISC parece tener un papel en la degradación del ADN durante el desarrollo del macronúcleo somático en los ciliados del género Tetrahymena . Es similar al control epigenético de la formación de la heterocromatina y se lo considera una defensa contra elementos genéticos invasores. [27]
De manera similar a la formación de heterocromatina en S. pombe y Arabidopsis , una proteína de Tetrahymena relacionada con la familia Argonaute, Twi1p, cataliza la eliminación del ADN de secuencias diana conocidas como secuencias de eliminación interna (IES). Mediante el uso de metiltransferasas y proteínas de cromodominio, las IES se heterocromatizan y se eliminan del ADN. [27]
Proteínas asociadas a RISC
La estructura completa de RISC aún no se ha resuelto. Muchos estudios han informado sobre una variedad de tamaños y componentes para RISC, pero no se sabe con certeza si esto se debe a que existen varios complejos RISC o a las diferentes fuentes que utilizan los distintos estudios. [28]
Ago, Argonaute; Dcr, Dicer; Dmp68, ortólogo de D. melanogaster de la p68 ARN desenrollasa de mamíferos; eIF2C1, factor de iniciación de la traducción eucariota 2C1; eIF2C2, factor de iniciación de la traducción eucariota 2C2; Fmr1/Fxr, ortólogo de D. melanogaster de la proteína de retraso mental del cromosoma X frágil; miRNP, complejo miRNA-proteína; NR, no informado; Tsn, nucleasa estafilocócica-Tudor; Vig, gen intrónico vasa.
Una proteína argonauta de longitud completa de la especie de arquea Pyrococcus furiosus .
De todas formas, es evidente que las proteínas Argonaute están presentes y son esenciales para la función. Además, se han obtenido datos sobre algunas de las proteínas clave (además de Argonaute) dentro del complejo, que permiten que RISC lleve a cabo su función.
Proteínas argonautas
Las proteínas Argonautas son una familia de proteínas que se encuentran en procariotas y eucariotas. Su función en procariotas es desconocida, pero en eucariotas son responsables de la interferencia del ARN. [43] Hay ocho miembros de la familia en los Argonautas humanos, de los cuales solo Argonauta 2 está involucrado exclusivamente en la escisión dirigida del ARN en RISC. [40]
El complejo de carga RISC permite cargar fragmentos de dsRNA (generados por Dicer) en Argonaute 2 (con la ayuda de TRBP) como parte de la vía de interferencia de ARN.
Complejo de carga RISC
El complejo de carga RISC (RLC) es la estructura esencial necesaria para cargar fragmentos de ARN bicatenario en el RISC con el fin de dirigirse al ARNm. El RLC está formado por Dicer, la proteína de unión al ARN de respuesta transactivadora ( TRBP ) y Argonaute 2.
Dicer es una endonucleasa ARNasa III que genera los fragmentos de ARNbc que se cargarán y que dirigen el ARNi.
TRBP es una proteína con tres dominios de unión a ARN bicatenario.
Argonaute 2 es una ARNasa y es el centro catalítico de RISC.
Dicer se asocia con TRBP y Argonaute 2 para facilitar la transferencia de los fragmentos de dsRNA generados por Dicer a Argonaute 2. [44] [45]
Investigaciones más recientes han demostrado que la helicasa A de ARN humana podría ayudar a facilitar la RLC. [46]
Otras proteínas
Los miembros recientemente identificados de RISC son SND1 y MTDH . [47] SND1 y MTDH son oncogenes y regulan la expresión de varios genes. [48]
Ago, Argonaute; Dcr, Dicer; Dmp68, ortólogo de D. melanogaster de la p68 ARN desenrollasa de mamíferos; eIF2C1, factor de iniciación de la traducción eucariota 2C1; eIF2C2, factor de iniciación de la traducción eucariota 2C2; Fmr1/Fxr, ortólogo de D. melanogaster de la proteína de retraso mental del cromosoma X frágil; Tsn, nucleasa estafilocócica de Tudor; Vig, gen intrónico de los vasos.
Unión del ARNm
Diagrama de la actividad RISC con miRNAs
Todavía no está claro cómo el complejo RISC activado localiza los objetivos del ARNm en la célula, aunque se ha demostrado que el proceso puede ocurrir en situaciones fuera de la traducción de proteínas en curso del ARNm. [50]
Los miRNA expresados endógenamente en metazoos no suelen ser perfectamente complementarios a una gran cantidad de genes y, por lo tanto, modulan la expresión a través de la represión traduccional. [51] [52] Sin embargo, en plantas , el proceso tiene una especificidad mucho mayor para el ARNm diana y, por lo general, cada miRNA solo se une a un ARNm. Una mayor especificidad significa que es más probable que se produzca la degradación del ARNm. [53]
^ abcdef Pratt AJ, MacRae IJ (2009). "El complejo silenciador inducido por ARN: una máquina versátil de silenciamiento de genes". Journal of Biological Chemistry . 284 (27): 17897–17901. doi : 10.1074/jbc.R900012200 . PMC 2709356 . PMID 19342379.
^ ab Filipowicz W, Bhattacharyya SN, Sonenber N (2008). "Mecanismos de regulación postranscripcional por microARN: ¿hay respuestas a la vista?". Nature Reviews Genetics . 9 (2): 102–114. doi :10.1038/nrg2290. PMID 18197166. S2CID 11824239.
^ ab Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, Mello CC (1998). "Interferencia genética potente y específica por ARN bicatenario en Caenorhabditis elegans". Nature . 391 (6669): 806–811. Bibcode :1998Natur.391..806F. doi :10.1038/35888. PMID 9486653. S2CID 4355692.
^ Watson, James D. (2008). Biología molecular del gen . San Francisco, CA: Cold Spring Harbor Laboratory Press. pp. 641–648. ISBN978-0-8053-9592-1.
^ abcd Hammond SM, Bernstein E , Beach D, Hannon GJ (2000). "Una nucleasa dirigida por ARN media el silenciamiento génico postranscripcional en células de Drosophila ". Nature . 404 (6775): 293–296. Bibcode :2000Natur.404..293H. doi :10.1038/35005107. PMID 10749213. S2CID 9091863.
^ Zamore PD, Tuschl T, Sharp PA, Bartel DP (2000). "RNAi: el ARN bicatenario dirige la escisión dependiente de ATP del ARNm en intervalos de 21 a 23 nucleótidos". Cell . 101 (1): 25–33. doi : 10.1016/S0092-8674(00)80620-0 . PMID 10778853.
^ Vermeulen A, Behlen L, Reynolds A, Wolfson A, Marshall W, Karpilow J, Khvorova A (2005). "Las contribuciones de la estructura del dsRNA a la especificidad y eficiencia de Dicer". ARN . 11 (5): 674–682. doi :10.1261/rna.7272305. PMC 1370754 . PMID 15811921.
^ Hutvagner, Gyorgy (2005). "Asimetría de ARN pequeño en RNAi: función en el ensamblaje de RISC y regulación génica". FEBS Letters . 579 (26): 5850–5857. doi : 10.1016/j.febslet.2005.08.071 . hdl : 10453/15313 . ISSN 1873-3468. PMID 16199039.
^ Schwarz DS, Hutvágner G, Du T, Xu Z, Aronin N, Zamore PD (2003). "Asimetría en el ensamblaje del complejo enzimático de ARNi". Cell . 115 (2): 199–208. doi : 10.1016/S0092-8674(03)00759-1 . PMID 14567917.
^ Khvorova A, Reynolds A, Jayasena SD (2003). "Los siRNA y miRNA funcionales exhiben sesgo de cadena". Cell . 115 (2): 209–216. doi : 10.1016/S0092-8674(03)00801-8 . PMID 14567918. S2CID 2500175.
^ ab Siomi H, Siomi MC (2009). "En el camino hacia la lectura del código de interferencia del ARN". Nature . 457 (7228): 396–404. Bibcode :2009Natur.457..396S. doi :10.1038/nature07754. PMID 19158785. S2CID 205215974.
^ Preall, Jonathan B.; Sontheimer, Erik J. (18 de noviembre de 2005). "RNAi: RISC se carga". Cell . 123 (4): 543–545. doi : 10.1016/j.cell.2005.11.006 . ISSN 0092-8674. PMID 16286001.
^ "Descripción general de la interferencia del ARN | Abcam" www.abcam.com . Consultado el 7 de marzo de 2021 .
^ Preall, Jonathan B.; He, Zhengying; Gorra, Jeffrey M.; Sontheimer, Erik J. (7 de marzo de 2006). "La selección de cadenas cortas de ARN interferente es independiente de la polaridad de procesamiento de dsRNA durante la interrelación del ARN en Drosophila". Current Biology . 16 (5): 530–535. Bibcode :2006CBio...16..530P. doi : 10.1016/j.cub.2006.01.061 . ISSN 0960-9822. PMID 16527750.
^ Gregory RI, Chendrimada TP, Cooch N, Shiekhattar R (2005). "El RISC humano acopla la biogénesis de microARN y el silenciamiento génico postranscripcional". Cell . 123 (4): 631–640. doi : 10.1016/j.cell.2005.10.022 . PMID 16271387.
^ Santhekadur, Prasanna K.; Kumar, Divya P. (1 de junio de 2020). "Ensamblaje de RISC y regulación génica postranscripcional en el carcinoma hepatocelular". Genes & Diseases . 7 (2): 199–204. doi : 10.1016/j.gendis.2019.09.009 . ISSN 2352-3042. PMC 7083748 . PMID 32215289.
^ ab Wakiyama M, Takimoto K, Ohara O, Yokoyama S (2007). "Deadenilación del ARNm mediada por microARN Let-7 y represión de la traducción en un sistema libre de células de mamíferos". Genes & Development . 21 (15): 1857–1862. doi :10.1101/gad.1566707. PMC 1935024 . PMID 17671087.
^ ab ORBAN, TAMAS I.; IZAURRALDE, ELISA (abril de 2005). "La desintegración de los ARNm dirigidos por RISC requiere XRN1, el complejo Ski y el exosoma". ARN . 11 (4): 459–469. doi :10.1261/rna.7231505. ISSN 1355-8382. PMC 1370735 . PMID 15703439.
^ Sen GL, Blau HM (2005). "Argonaute2/RISC reside en sitios de desintegración del ARNm de mamíferos conocidos como cuerpos citoplasmáticos". Nature Cell Biology . 7 (6): 633–636. doi :10.1038/ncb1265. PMID 15908945. S2CID 6085169.
^ Chendrimada TP, Finn KJ, Ji X, Baillat D, Gregory RI, Liebhaber SA, Pasquinelli AE, Shiekhattar R (2007). "Silenciamiento de microARN mediante reclutamiento de eIF6 por RISC". Nature . 447 (7146): 823–828. Bibcode :2007Natur.447..823C. doi :10.1038/nature05841. PMID 17507929. S2CID 4413327.
^ Petersen CP, Bordeleau ME, Pelletier J, Sharp PA (2006). "Los ARN cortos reprimen la traducción después de la iniciación en células de mamíferos". Molecular Cell . 21 (4): 533–542. doi : 10.1016/j.molcel.2006.01.031 . PMID 16483934.
^ Maroney PA, Yu Y, Fisher J, Nilsen TW (2006). "Evidencia de que los microARN están asociados con la traducción de ARN mensajeros en células humanas". Nature Structural & Molecular Biology . 13 (12): 1102–1107. doi :10.1038/nsmb1174. PMID 17128271. S2CID 19106463.
^ Verdel A, Jia S, Gerber S, Sugiyama T, Gygi S, Grewal SI, Moazed D (2004). "Orientación de la heterocromatina mediada por ARNi por el complejo RITS". Science . 303 (5658): 672–676. Bibcode :2004Sci...303..672V. doi :10.1126/science.1093686. PMC 3244756 . PMID 14704433.
^ Verdel A, Jia S, Gerber S, Sugiyama T, Gygi S, Grewal SI, Moazed D (2004). "RITS actúa en cis para promover la transcripción mediada por interferencia de ARN y el silenciamiento postranscripcional". Nature Genetics . 36 (11): 1174–1180. doi : 10.1038/ng1452 . PMID 15475954.
^ Shimada, Yukiko; Mohn, Fabio; Bühler, Marc (1 de diciembre de 2016). "El complejo silenciador transcripcional inducido por ARN se dirige exclusivamente a la cromatina a través de la interacción con transcripciones nacientes". Genes & Development . 30 (23): 2571–2580. doi :10.1101/gad.292599.116. ISSN 0890-9369. PMC 5204350 . PMID 27941123.
^ Sugiyama T, Cam H, Verdel A, Moazed D, Grewal SI (2005). "La ARN polimerasa dependiente de ARN es un componente esencial de un acoplamiento de bucle autoimpuesto del ensamblaje de la heterocromatina a la producción de ARNi". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 102 (1): 152–157. doi : 10.1073/pnas.0407641102 . PMC 544066 . PMID 15615848.
^ abc Mochizuki K, Gorovsky MA (2004). "Pequeños ARN en la disposición del genoma en Tetrahymena ". Current Opinion in Genetics & Development . 14 (2): 181–187. doi :10.1016/j.gde.2004.01.004. PMID 15196465.
^ abc Sontheimer EJ (2005). "Ensamblaje y función de complejos silenciadores de ARN". Nature Reviews Molecular Cell Biology . 6 (2): 127–138. doi :10.1038/nrm1568. PMID 15654322. S2CID 27294007.
^ ab Liu Q, Rand TA, Kalidas S, Du F, Kim HE, Smith DP, Wang X (2003). "R2D2, un puente entre los pasos de iniciación y efector de la vía de RNAi de Drosophila ". Science . 301 (5641): 1921–1925. Bibcode :2003Sci...301.1921L. doi :10.1126/science.1088710. PMID 14512631. S2CID 41436233.
^ abcdefghij Pham JW, Pellio JL, Lee YS, Carthew RW, Sontheimer EJ (2004). "Un complejo 80S dependiente de Dicer-2 escinde ARNm específicos durante la interrelación del ARN en Drosophila". Cell . 117 (1): 83–94. doi : 10.1016/S0092-8674(04)00258-2 . PMID 15066284.
^ abcd Tomari Y, Du T, Haley B, Schwarz DS, Bennett R, Cook HA, Koppetsch BS, Theurkauf WE, Zamore PD (2004). "Defectos de ensamblaje de RISC en el mutante de ARNi de Drosophila armitage". Cell . 116 (6): 831–841. doi : 10.1016/S0092-8674(04)00218-1 . PMID 15035985.
^ abc Hammond SM, Boettcher S, Caudy AA, Kobayashi R, Hannon GJ (2001). "Argonaute2, un vínculo entre los análisis genéticos y bioquímicos del ARNi". Science . 293 (5532): 1146–1150. doi :10.1126/science.1064023. PMID 11498593. S2CID 5271290.
^ abc Caudy AA, Myers M, Hannon GJ, Hammond SM (2002). "La proteína relacionada con el cromosoma X frágil y la VIG se asocian con la maquinaria de interferencia del ARN". Genes & Development . 16 (19): 2491–2496. doi :10.1101/gad.1025202. PMC 187452 . PMID 12368260.
^ ab Caudy AA, Ketting RF, Hammond SM, Denli AM, Bathoorn AM, Tops BB, Silva JM, Myers MM, Hannon GJ, Plasterk RH (2003). "Un homólogo de nucleasa microcócica en complejos efectores de ARNi". Nature . 425 (6956): 411–414. Bibcode :2003Natur.425..411C. doi :10.1038/nature01956. PMID 14508492. S2CID 4410688.
^ ab Rand TA, Ginalski K, Grishin NV, Wang X (2004). "Identificación bioquímica de Argonaute 2 como la única proteína necesaria para la actividad del complejo silenciador inducido por ARN". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 101 (40): 14385–14389. Bibcode :2004PNAS..10114385R. doi : 10.1073/pnas.0405913101 . PMC 521941 . PMID 15452342.
^ abcde Ishizuka A, Siomi MC, Siomi H (2002). "Una proteína X frágil de Drosophila interactúa con componentes de ARNi y proteínas ribosomales". Genes & Development . 16 (19): 2497–2508. doi :10.1101/gad.1022002. PMC 187455 . PMID 12368261.
^ abc Martinez J, Patkaniowska A, Urlaub H, Luhrmann R, Tuschl T (2002). "Los siRNA antisentido monocatenarios guían la escisión del ARN diana en la interrelación del ARN". Cell . 110 (5): 563–574. doi :10.1016/S0092-8674(02)00908-X. hdl : 11858/00-001M-0000-0012-F2FD-2 . PMID 12230974. S2CID 10616773.
^ ab Liu J, Carmell MA, Rivas FV, Marsden CG, Thomson JM, Song JJ, Hammond SM, Joshua-Tor L, Hannon GJ (2004). "Argonaute2 es el motor catalítico del ARNi en mamíferos". Science . 305 (5689): 1437–1441. Bibcode :2004Sci...305.1437L. doi : 10.1126/science.1102513 . PMID 15284456. S2CID 2778088.
^ Martinez J, Tuschl T (2004). "RISC es una ARN endonucleasa productora de fosfomonoésteres 5'". Genes & Development . 18 (9): 975–980. doi :10.1101/gad.1187904. PMC 406288 . PMID 15105377.
^ abc Meister G, Landthaler M, Patkaniowska A, Dorsett Y, Teng G, Tuschl T (2004). "El Argonaute2 humano media la escisión del ARN dirigida por los miRNA y los siRNA". Molecular Cell . 15 (2): 1403–1408. doi : 10.1016/j.molcel.2004.07.007 . PMID 15260970.
^ abc Mourelatos Z, Dostie J, Paushkin S, Sharma A, Charroux B, Abel L, Rappsilber J , Mann M, Dreyfuss G (2002). "miRNPs: una nueva clase de ribonucleoproteínas que contienen numerosos microARN". Genes & Development . 16 (6): 720–728. doi :10.1101/gad.974702. PMC 155365 . PMID 11914277.
^ abcd Hutvágner G, Zamore PD (2002). "Un microARN en un complejo enzimático de ARNi de recambio múltiple". Science . 297 (5589): 2056–2060. Bibcode :2002Sci...297.2056H. doi :10.1126/science.1073827. PMID 12154197. S2CID 16969059.
^ Hall TM (2005). "Estructura y función de las proteínas Argonaute". Cell . 13 (10): 1403–1408. doi : 10.1016/j.str.2005.08.005 . PMID 16216572.
^ Chendrimada TP, Gregory RI, Kumaraswamy E, Norman J, Cooch N, Nishikura K, Shiekhatter R (2005). "TRBP recluta el complejo Dicer a Ago2 para el procesamiento de microARN y el silenciamiento génico". Nature . 436 (7051): 740–744. Bibcode :2005Natur.436..740C. doi :10.1038/nature03868. PMC 2944926 . PMID 15973356.
^ Wang HW, Noland C, Siridechadilok B, Taylor DW, Ma E, Felderer K, Doudna JA, Nogales E (2009). "Información estructural sobre el procesamiento del ARN por el complejo de carga RISC humano". Nature Structural & Molecular Biology . 16 (11): 1148–1153. doi :10.1038/nsmb.1673. PMC 2845538 . PMID 19820710.
^ Fu Q, Yuan YA (2013). "Información estructural sobre el ensamblaje de RISC facilitado por los dominios de unión a dsRNA de las hélices A de ARN humano (DHX9)". Investigación de ácidos nucleicos . 41 (5): 3457–3470. doi :10.1093/nar/gkt042. PMC 3597700 . PMID 23361462.
^ Yoo BK, Santhekadur PK, Gredler R, Chen D, Emdad L, Bhutia S, Pannell L, Fisher PB, Sarkar D (2011). "El aumento de la actividad del complejo silenciador inducido por ARN (RISC) contribuye al carcinoma hepatocelular". Hepatología . 53 (5): 1538–1548. doi :10.1002/hep.24216. PMC 3081619 . PMID 21520169.
^ Yoo BK, Emdad L, Lee SG, Su Z, Santhekadur P, Chen D, Gredler R, Fisher PB, Sarkar D (2011). "Gen elevado de astrocitos (AEG-1): un regulador multifuncional de la fisiología normal y anormal". Farmacología y terapéutica . 130 (1): 1–8. doi :10.1016/j.pharmthera.2011.01.008. PMC 3043119 . PMID 21256156.
^ Djikeng A, Shi H, Tschudi C, Shen S, Ullu E (2003). "Se ha encontrado una ribonucleoproteína de ARNi asociada a polirribosomas en Trypanosoma brucei". ARN . 9 (7): 802–808. doi :10.1261/rna.5270203. PMC 1370447 . PMID 12810914.
^ Sen GL, Wehrman TS, Blau HM (2005). "La traducción del ARNm no es un requisito previo para la escisión del ARNm mediada por ARN interferentes pequeños". Differentiation . 73 (6): 287–293. doi :10.1111/j.1432-0436.2005.00029.x. PMID 16138829.
^ Saumet A, Lecellier CH (2006). "Silenciamiento del ARN antiviral: ¿nos parecemos a las plantas?". Retrovirology . 3 : 3. doi : 10.1186/1742-4690-3-3 . PMC 1363733 . PMID 16409629.
^ Bartel DP (2009). "MicroRNAs: reconocimiento de dianas y funciones reguladoras". Cell . 136 (2): 215–233. doi :10.1016/j.cell.2009.01.002. PMC 3794896 . PMID 19167326.
^ Jones-Rhoades MW, Bartel DP, Bartel B (2006). "MicroRNAs and their regulator roles in plants" (MicroARN y sus funciones reguladoras en plantas). Revisión anual de biología vegetal . 57 : 19–53. doi :10.1146/annurev.arplant.57.032905.105218. PMID 16669754.
Lectura adicional
Sontheimer, EJ (2005). "Ensamblaje y función de complejos silenciadores de ARN". Nature Reviews Molecular Cell Biology . 6 (2): 127–138. doi :10.1038/nrm1568. PMID 15654322. S2CID 27294007.
Fu Q, Yuan YA (marzo de 2013). "Información estructural sobre el ensamblaje de RISC facilitado por los dominios de unión a dsRNA de la helicasa A de ARN humana (DHX9)". Nucleic Acids Research . 41 (5): 3457–70. doi :10.1093/nar/gkt042. PMC 3597700 . PMID 23361462.
Schwarz DS, Tomari Y, Zamore PD (2004). "El complejo de silenciamiento inducido por ARN es una endonucleasa dependiente de Mg2+". Current Biology . 14 (9): 787–91. Bibcode :2004CBio...14..787S. doi : 10.1016/j.cub.2004.03.008 . PMID 15120070.
.jpg/1280px-Drosophila_melanogaster_-_side_(aka).jpg)