El circuito cortico-basal ganglio-tálamo-cortical ( circuito CBGTC ) es un sistema de circuitos neuronales en el cerebro . El circuito involucra conexiones entre la corteza , los ganglios basales , el tálamo y de regreso a la corteza. Es de particular relevancia para los trastornos del movimiento hipercinéticos e hipocinéticos , como la enfermedad de Parkinson y la enfermedad de Huntington , [1] así como para los trastornos mentales de control, como el trastorno por déficit de atención con hiperactividad (TDAH), [2] el trastorno obsesivo-compulsivo (TOC), [3] y el síndrome de Tourette . [4]
El bucle CBGTC consta principalmente de proyecciones dopaminérgicas moduladoras de la pars compacta de la sustancia negra y el área tegmental ventral , así como proyecciones glutamatérgicas excitatorias de la corteza al cuerpo estriado , donde estas proyecciones forman sinapsis con vías excitatorias e inhibidoras que retransmiten de vuelta a la corteza. El bucle se propuso originalmente como parte de un modelo de los ganglios basales llamado modelo de procesamiento paralelo , que ha sido criticado y modificado en otro modelo llamado modelo de centro envolvente . [5]
Los esquemas de organización actuales caracterizan las interacciones cortico-ganglios basales como un procesamiento paralelo segregado, lo que significa que hay poca convergencia de áreas corticales distintas en los ganglios basales. Se cree que esto explica la funcionalidad organizada topográficamente del cuerpo estriado. [4] El cuerpo estriado está organizado en un eje rostro-caudal, con el putamen rostral y el caudado cumpliendo funciones asociativas y cognitivas y las áreas caudales cumpliendo funciones sensoriomotoras. [6] A veces, cuando el cuerpo estriado es el objetivo expresado, el bucle se denomina bucle cortico-estriatal-tálamo-cortical . [7]
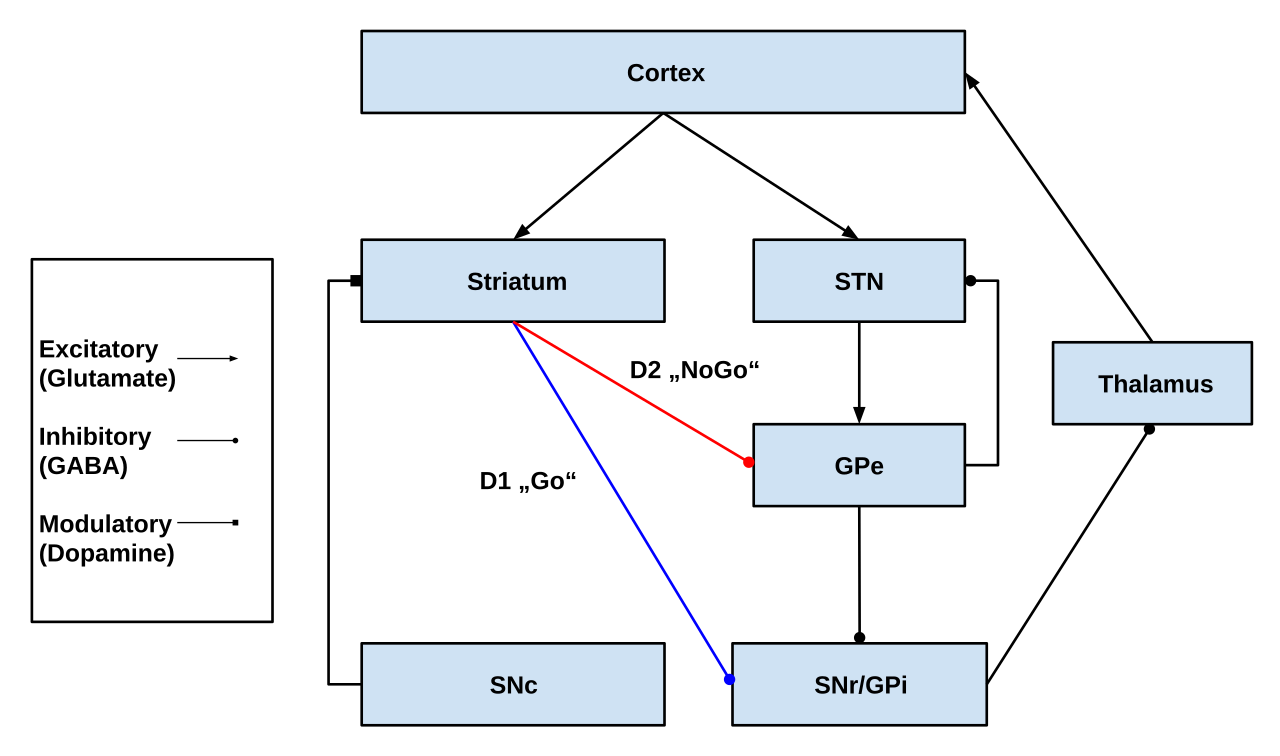
Las dos estructuras de entrada principales del circuito son el cuerpo estriado y el núcleo subtalámico (STN). El cuerpo estriado recibe entradas tanto de la corteza como de la pars compacta de la sustancia negra (SNc), mientras que el STN solo recibe entradas corticales.
Del cuerpo estriado surgen dos vías: una de ellas, denominada vía indirecta (o vía NoGo) , es inhibidora y se proyecta hacia el globo pálido externo (GPe) y lo inhibe, lo que da lugar a la desinhibición del globo pálido interno (GPi), lo que conduce a la inhibición del tálamo. Esta vía también, como resultado de la inhibición del GPe, desinhibe el núcleo subtalámico, lo que da lugar a la excitación del GPi y, por lo tanto, a la inhibición del tálamo.
La segunda vía, llamada vía directa (o Go) y es excitatoria. Esta vía inhibe el GPi, lo que da como resultado la desinhibición del tálamo. La vía directa consiste principalmente en conexiones monosinápticas impulsadas por el receptor de dopamina D1 , el receptor de adenosina A1 y el receptor muscarínico de acetilcolina M4 , mientras que la vía indirecta se basa en conexiones impulsadas por el receptor de dopamina D2 , el receptor de adenosina A2A y el receptor muscarínico de acetilcolina M1 . [1] [8]
Los circuitos paralelos de CBGTC se han dividido según las funciones de las regiones corticales asociadas. Un esquema implica la división en circuitos límbicos y motores, donde los circuitos motores contienen vías indirectas y directas, que a su vez están interconectadas con el circuito límbico que se proyecta hacia el cuerpo estriado ventral. [9] El circuito también se ha dividido en circuitos límbicos, asociativos, oculomotores y motores [4] para explicar el papel de la dopamina en los ganglios basales sobre los estados motivacionales. [10] Se ha descrito una división de cinco circuitos basada en objetivos corticales primarios de la siguiente manera: [11]
Un problema identificado con la anatomía actual del circuito es que el retraso temporal entre las vías directas e indirectas debería hacer que este circuito no funcione. Para superar esto, la hipótesis de centro envolvente postula que una vía hiperdirecta desde la corteza inhibiría otras entradas además de una entrada cortical enfocada. Sin embargo, el momento de la actividad de los ganglios basales y el momento de las extremidades, así como los estudios de lesiones, no respaldan esta hipótesis [12].
Se han propuesto dos modelos para explicar cómo se seleccionan las acciones en los ganglios basales. El modelo actor-crítico sugiere que las acciones son generadas y evaluadas por un "crítico" en el cuerpo estriado ventral, mientras que las acciones son llevadas a cabo por un "actor" en el cuerpo estriado dorsal. Otro modelo propone que los ganglios basales actúan como un mecanismo de selección, donde las acciones son generadas en la corteza y son seleccionadas en función del contexto por los ganglios basales. [13] El circuito CBGTC también está involucrado en el descuento de recompensas, con un aumento de la activación con una recompensa inesperada o mayor a la esperada. [2] Una revisión apoyó la idea de que la corteza estaba involucrada en el aprendizaje de acciones independientemente de su resultado, mientras que los ganglios basales estaban involucrados en la selección de acciones apropiadas basadas en el aprendizaje asociativo de prueba y error basado en recompensas. [14]
El circuito CBGTC se ha relacionado con muchas enfermedades. Por ejemplo, en la enfermedad de Parkinson , se cree que la degeneración de las neuronas dopaminérgicas que conduce a una actividad reducida de la vía excitatoria produce hipocinesia [15], y en la enfermedad de Huntington , se cree que la degeneración de las neuronas GABAérgicas que impulsan la vía inhibidora produce movimientos corporales espasmódicos [2] . La degeneración simultánea de las proyecciones límbicas junto con las proyecciones motoras puede producir muchos de los síntomas psiquiátricos de estas enfermedades principalmente motoras [9] . En el TOC, el circuito puede ser disfuncional, con un desequilibrio entre las vías indirectas y directas que da lugar a pensamientos no deseados que se "quedan estancados". [3] En el TDAH, se cree que la disminución de la señalización dopaminérgica tónica que da lugar a un descuento excesivo de las recompensas retrasadas produce una disminución de la atención [2] .
El bucle CBGTC se ha estudiado en relación con la conciencia , la selección de acciones, en relación con otros circuitos y en el contexto de la memoria y la cognición. [16] [17] El modelo del bucle CBGTC ha sido criticado por simplificarse demasiado y aplicarse de forma demasiado rígida, dada la evidencia de superposición anatómica y funcional e interacciones entre las vías directas e indirectas. [18] El bucle también se ha investigado en el contexto de la estimulación cerebral profunda . [16] A partir de 2013, hubo un intenso debate con respecto a la división del circuito, las interacciones de las vías, el número de vías y la anatomía general. [17]
{{cite journal}}: Requiere citar revista |journal=( ayuda )