Molécula de proteína que recibe señales para una célula.
Un ejemplo de receptores de membrana .
Ligandos, ubicados fuera de la célula.
Los ligandos se conectan a proteínas receptoras específicas según la forma del sitio activo de la proteína.
El receptor libera un mensajero una vez que el ligando se ha conectado al receptor.
En bioquímica y farmacología , los receptores son estructuras químicas, compuestas de proteínas , que reciben y transducen señales que pueden integrarse en sistemas biológicos. [1] Estas señales suelen ser mensajeros químicos [nb 1] que se unen a un receptor y producen respuestas fisiológicas , como cambios en la actividad eléctrica de una célula . Por ejemplo, el GABA , un neurotransmisor inhibidor , inhibe la actividad eléctrica de las neuronas uniéndose a los receptores GABA A. [2] Hay tres formas principales en que se puede clasificar la acción del receptor: retransmisión de señal, amplificación o integración. [3] La retransmisión envía la señal, la amplificación aumenta el efecto de un solo ligando y la integración permite que la señal se incorpore a otra vía bioquímica. [3]
Las proteínas receptoras se pueden clasificar según su ubicación. Los receptores de la superficie celular , también conocidos como receptores transmembrana, incluyen canales iónicos activados por ligando , receptores acoplados a proteína G y receptores hormonales ligados a enzimas . [1] Los receptores intracelulares son aquellos que se encuentran dentro de la célula e incluyen receptores citoplasmáticos y receptores nucleares . [1] Una molécula que se une a un receptor se llama ligando y puede ser una proteína, un péptido (proteína corta) u otra molécula pequeña , como un neurotransmisor , una hormona , un fármaco, una toxina, un ion de calcio o partes del exterior. de un virus o microbio. Una sustancia producida endógenamente que se une a un receptor particular se denomina ligando endógeno. Por ejemplo, el ligando endógeno para el receptor nicotínico de acetilcolina es la acetilcolina , pero también puede activarse con nicotina [4] [5] y bloquearse con curare . [6] Los receptores de un tipo particular están vinculados a vías bioquímicas celulares específicas que corresponden a la señal. Si bien en la mayoría de las células se encuentran numerosos receptores, cada receptor solo se unirá a ligandos de una estructura particular. Esto se ha comparado de manera análoga a cómo las cerraduras sólo aceptan llaves con formas específicas . Cuando un ligando se une a un receptor correspondiente, activa o inhibe la vía bioquímica asociada al receptor, que también puede ser altamente especializada.
Las estructuras de los receptores son muy diversas e incluyen, entre otras, las siguientes categorías principales:
Tipo 1: canales iónicos activados por ligando (receptores ionotrópicos): estos receptores suelen ser el objetivo de neurotransmisores rápidos como la acetilcolina (nicotínico) y el GABA ; La activación de estos receptores produce cambios en el movimiento de los iones a través de una membrana. Tienen una estructura heteromérica en la que cada subunidad consta del dominio de unión al ligando extracelular y un dominio transmembrana que incluye cuatro hélices alfa transmembrana . Las cavidades de unión al ligando están ubicadas en la interfaz entre las subunidades.
Tipo 2: Receptores acoplados a proteína G (receptores metabotrópicos): esta es la familia más grande de receptores e incluye receptores para varias hormonas y transmisores lentos, por ejemplo, dopamina y glutamato metabotrópico. Están compuestos por siete hélices alfa transmembrana. Los bucles que conectan las hélices alfa forman dominios extracelulares e intracelulares. El sitio de unión para ligandos peptídicos más grandes generalmente se ubica en el dominio extracelular, mientras que el sitio de unión para ligandos no peptídicos más pequeños a menudo se ubica entre las siete hélices alfa y un bucle extracelular. [7] Los receptores antes mencionados están acoplados a diferentes sistemas efectores intracelulares a través de proteínas G. [8] Las proteínas G son heterotrímeros formados por 3 subunidades: α (alfa), β (beta) y γ (gamma). En el estado inactivo, las tres subunidades se asocian y la subunidad α se une al PIB. [9] La activación de la proteína G provoca un cambio conformacional, que conduce al intercambio de GDP por GTP. La unión de GTP a la subunidad α provoca la disociación de las subunidades β y γ. [10] Además, las tres subunidades, α, β y γ, tienen cuatro clases principales adicionales basadas en su secuencia primaria. Estos incluyen G s , G i , G q y G 12 . [11]
Tipo 3: Receptores ligados a quinasa y relacionados (ver " Receptor tirosina quinasa " y " Receptor ligado a enzima "): están compuestos por un dominio extracelular que contiene el sitio de unión del ligando y un dominio intracelular, a menudo con función enzimática, unido por una única hélice alfa transmembrana. El receptor de insulina es un ejemplo.
Tipo 4: Receptores nucleares : si bien se les llama receptores nucleares, en realidad están ubicados en el citoplasma y migran al núcleo después de unirse con sus ligandos. Están compuestos por una región de unión al ligando C-terminal , un dominio de unión al ADN central (DBD) y un dominio N-terminal que contiene la región AF1 (función de activación 1). La región central tiene dos dedos de zinc que se encargan de reconocer las secuencias de ADN específicas de este receptor. El extremo N interactúa con otros factores de transcripción celular de manera independiente del ligando; y, dependiendo de estas interacciones, puede modificar la unión/actividad del receptor. Los receptores de esteroides y de hormonas tiroideas son ejemplos de tales receptores. [12]
Ligand binding is an equilibrium process. Ligands bind to receptors and dissociate from them according to the law of mass action in the following equation, for a ligand L and receptor, R. The brackets around chemical species denote their concentrations.
One measure of how well a molecule fits a receptor is its binding affinity, which is inversely related to the dissociation constantKd. A good fit corresponds with high affinity and low Kd. The final biological response (e.g. second messenger cascade, muscle-contraction), is only achieved after a significant number of receptors are activated.
Affinity is a measure of the tendency of a ligand to bind to its receptor. Efficacy is the measure of the bound ligand to activate its receptor.
Agonists versus antagonists
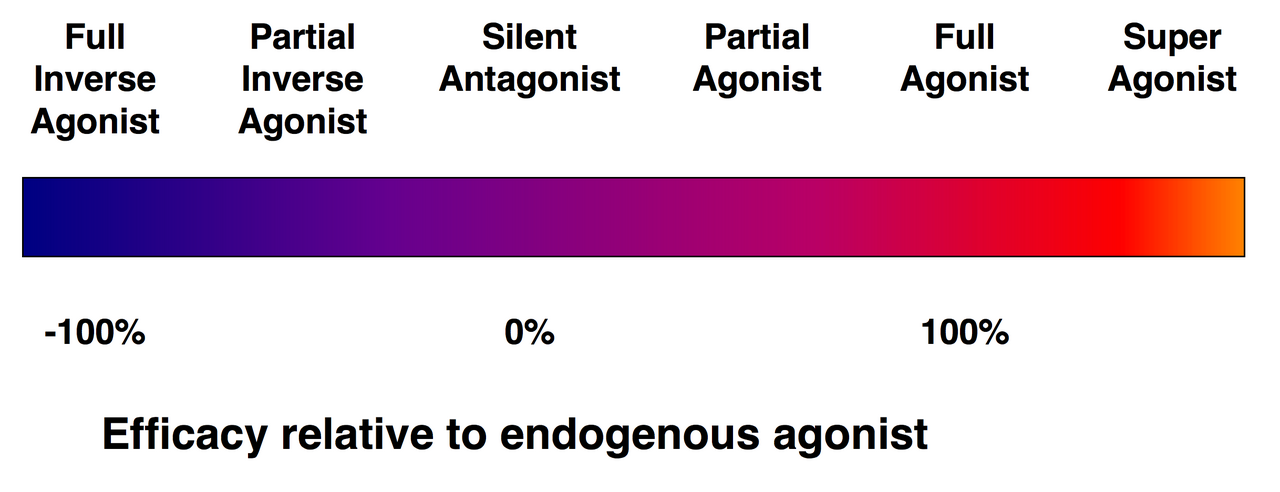
Efficacy spectrum of receptor ligands.
Not every ligand that binds to a receptor also activates that receptor. The following classes of ligands exist:
(Full) agonists are able to activate the receptor and result in a strong biological response. The natural endogenous ligand with the greatest efficacy for a given receptor is by definition a full agonist (100% efficacy).
Partial agonists do not activate receptors with maximal efficacy, even with maximal binding, causing partial responses compared to those of full agonists (efficacy between 0 and 100%).
Antagonists bind to receptors but do not activate them. This results in a receptor blockade, inhibiting the binding of agonists and inverse agonists. Receptor antagonists can be competitive (or reversible), and compete with the agonist for the receptor, or they can be irreversible antagonists that form covalent bonds (or extremely high affinity non-covalent bonds) with the receptor and completely block it. The proton pump inhibitor omeprazole is an example of an irreversible antagonist. The effects of irreversible antagonism can only be reversed by synthesis of new receptors.
Inverse agonists reduce the activity of receptors by inhibiting their constitutive activity (negative efficacy).
Moduladores alostéricos : no se unen al sitio de unión del agonista del receptor sino a sitios de unión alostéricos específicos, a través de los cuales modifican el efecto del agonista. Por ejemplo, las benzodiazepinas (BZD) se unen al sitio BZD en el receptor GABA A y potencian el efecto del GABA endógeno.
Tenga en cuenta que la idea de agonismo y antagonismo del receptor solo se refiere a la interacción entre receptores y ligandos y no a sus efectos biológicos.
Actividad constitutiva
Se dice que un receptor que es capaz de producir una respuesta biológica en ausencia de un ligando unido muestra "actividad constitutiva". [13] La actividad constitutiva de un receptor puede ser bloqueada por un agonista inverso . Los medicamentos contra la obesidad rimonabant y taranabant son agonistas inversos del receptor cannabinoide CB1 y, aunque produjeron una pérdida de peso significativa, ambos fueron retirados debido a una alta incidencia de depresión y ansiedad, que se cree que están relacionadas con la inhibición de la actividad constitutiva de El receptor cannabinoide.
El receptor GABA A tiene actividad constitutiva y conduce cierta corriente basal en ausencia de un agonista. Esto permite que la beta carbolina actúe como un agonista inverso y reduzca la corriente por debajo de los niveles basales.
Las mutaciones en los receptores que dan como resultado una mayor actividad constitutiva son la base de algunas enfermedades hereditarias, como la pubertad precoz (debido a mutaciones en los receptores de la hormona luteinizante) y el hipertiroidismo (debido a mutaciones en los receptores de la hormona estimulante de la tiroides).
Teorías de la interacción fármaco-receptor.
Ocupación
Las primeras formas de la teoría farmacológica de los receptores afirmaban que el efecto de un fármaco es directamente proporcional al número de receptores ocupados. [14] Además, el efecto de un fármaco cesa cuando el complejo fármaco-receptor se disocia.
Ariëns y Stephenson introdujeron los términos "afinidad" y "eficacia" para describir la acción de ligandos unidos a receptores. [15] [16]
Afinidad : Capacidad de un fármaco para combinarse con un receptor para crear un complejo fármaco-receptor.
Eficacia : Capacidad de un fármaco para iniciar una respuesta después de la formación del complejo fármaco-receptor.
Tasa
En contraste con la Teoría de la Ocupación aceptada , la Teoría de la Tasa propone que la activación de los receptores es directamente proporcional al número total de encuentros de una droga con sus receptores por unidad de tiempo. La actividad farmacológica es directamente proporcional a las tasas de disociación y asociación, no al número de receptores ocupados: [17]
Agonista: Fármaco con una rápida asociación y una rápida disociación.
Agonista parcial: fármaco con una asociación intermedia y una disociación intermedia.
Antagonista: fármaco de asociación rápida y disociación lenta.
Ajuste inducido
Cuando un fármaco se acerca a un receptor, el receptor altera la conformación de su sitio de unión para producir un complejo fármaco-receptor.
Receptores de repuesto
En algunos sistemas receptores (p. ej., acetilcolina en la unión neuromuscular del músculo liso), los agonistas pueden provocar una respuesta máxima a niveles muy bajos de ocupación del receptor (<1%). Por tanto, ese sistema tiene receptores de repuesto o una reserva de receptores. Esta disposición produce una economía de producción y liberación de neurotransmisores. [12]
Secuestro de receptores (internalización) [18] , por ejemplo en el caso de receptores hormonales.
Ejemplos y ligandos
Los ligandos de los receptores son tan diversos como sus receptores. Los GPCR (7TM) son una familia particularmente amplia, con al menos 810 miembros. También existen LGIC para al menos una docena de ligandos endógenos y muchos más receptores posibles a través de diferentes composiciones de subunidades. Algunos ejemplos comunes de ligandos y receptores incluyen: [19]
Canales iónicos y receptores acoplados a proteína G.
En la siguiente tabla se muestran algunos ejemplos de receptores ionotrópicos (LGIC) y metabotrópicos (específicamente, GPCR). Los principales neurotransmisores son el glutamato y el GABA; Otros neurotransmisores son neuromoduladores . Esta lista no es de ninguna manera exhaustiva.
Receptores ligados a enzimas
Los receptores ligados a enzimas incluyen receptores tirosina quinasas (RTK), proteína quinasa específica de serina/treonina, como en la proteína morfogenética ósea y guanilato ciclasa, como en el receptor del factor natriurético auricular. De los RTK, se han identificado 20 clases, con 58 RTK diferentes como miembros. Algunos ejemplos se muestran a continuación:
Receptores intracelulares
Los receptores pueden clasificarse según su mecanismo o su posición en la célula. A continuación se muestran 4 ejemplos de LGIC intracelular:
Papel en la salud y la enfermedad.
En trastornos genéticos
Muchos trastornos genéticos implican defectos hereditarios en los genes receptores. A menudo, es difícil determinar si el receptor no funciona o si la hormona se produce en un nivel reducido; esto da lugar al grupo de trastornos endocrinos "pseudo-hipo-" , donde parece haber un nivel hormonal disminuido cuando en realidad es el receptor el que no responde suficientemente a la hormona.
^ a b C Guyton, Arthur C.; Salón, John E. (2016). Libro de texto de fisiología médica de Guyton y Hall. Filadelfia, PA: Elsevier Saunders. págs. 930–937. ISBN 9781455770052. OCLC 1027900365.
^ Mihic, S. Juan; Harris, R. Adron (1997). "GABA y el receptor GABAA". Mundo de la investigación y la salud del alcohol . 21 (2): 127-131. ISSN 0090-838X. PMC 6826832 . PMID 15704348.
^ ab Alberts B, Bray D, Hopkin K, Johnson A, Lewis J, Raff M, Roberts K, Walter P (2014). Biología celular esencial (Cuarta ed.). Nueva York, NY, Estados Unidos: Garland Science. pag. 534.ISBN _978-0-8153-4454-4.
^ Gotti, Cecilia; Marcos, Miguel. J.; Millar, Neil S.; Wonnacott, Susan (16 de septiembre de 2019). "Receptores nicotínicos de acetilcolina (versión 2019.4)". Guía IUPHAR/BPS de Farmacología CITE . 2019 (4). doi : 10.2218/gtopdb/F76/2019.4 . Consultado el 17 de noviembre de 2020 .
^ Malenka RC, Nestler EJ, Hyman SE (2009). "Capítulo 9: Sistema nervioso autónomo". En Sydor A, Brown RY (eds.). Neurofarmacología molecular: una base para la neurociencia clínica (2ª ed.). Nueva York: McGraw-Hill Medical. pag. 234.ISBN _9780071481274. La nicotina... es un alcaloide natural de la planta del tabaco. La lobelina es un alcaloide natural del tabaco indio. Ambos fármacos son agonistas [de] los receptores colinérgicos nicotínicos...
^ "Información sobre medicamentos Curare, profesional". Drogas.com . Consultado el 8 de diciembre de 2020 .
^ Congreve M, Marshall F (marzo de 2010). "El impacto de las estructuras GPCR en la farmacología y el diseño de fármacos basado en estructuras". Revista británica de farmacología . 159 (5): 986–96. doi :10.1111/j.1476-5381.2009.00476.x. PMC 2839258 . PMID 19912230.
^ Qin K, Dong C, Wu G, Lambert NA (agosto de 2011). "Preensamblaje en estado inactivo de receptores acoplados a G (q) y heterotrímeros de G (q)". Biología Química de la Naturaleza . 7 (10): 740–7. doi :10.1038/nchembio.642. PMC 3177959 . PMID 21873996.
^ Zubay, Geoffrey (1998). Bioquímica 4ª Ed . Dubuque, IA: Pub William C Brown. pag. 684.ISBN _0697219003.
^ Hamm, Heidi E.; Oldham, William M. (2008). "Activación de la proteína G heterotrimérica por receptores acoplados a proteína G". Reseñas de la naturaleza Biología celular molecular . Grupo Editorial Naturaleza. 9 (1): 60–71. doi :10.1038/nrm2299. PMID 18043707. S2CID 24267759.
^ ab Rang HP, Dale MM, Ritter JM, Flower RJ, Henderson G (2012). Farmacología de Rang & Dale (7ª ed.). Elsevier Churchill Livingstone. ISBN978-0-7020-3471-8.
^ Milligan G (diciembre de 2003). "Actividad constitutiva y agonistas inversos de los receptores acoplados a proteína G: una perspectiva actual". Farmacología molecular . 64 (6): 1271–6. doi : 10,1124/mol.64.6.1271. PMID 14645655. S2CID 2454589.
^ Sonó, HP (enero de 2006). "El concepto de receptor: la gran idea de la farmacología". Revista británica de farmacología . 147 (Suplemento 1): T9-16. doi : 10.1038/sj.bjp.0706457. PMC 1760743 . PMID 16402126.
^ Ariens EJ (septiembre de 1954). "Afinidad y actividad intrínseca en la teoría de la inhibición competitiva. I. Problemas y teoría". Archivos Internacionales de Farmacodinamia y Terapia . 99 (1): 32–49. PMID 13229418.
^ Stephenson RP (diciembre de 1956). "Una modificación de la teoría del receptor". Revista británica de farmacología y quimioterapia . 11 (4): 379–93. doi :10.1111/j.1476-5381.1956.tb00006.x. PMC 1510558 . PMID 13383117.
^ Silverman RB (2004). "3.2.C Teorías sobre las interacciones entre fármacos y receptores". La química orgánica del diseño y la acción de los fármacos (2ª ed.). Ámsterdam: Elsevier Academic Press. ISBN0-12-643732-7.
^ Boulay G, Chrétien L, Richard DE, Guillemette G (noviembre de 1994). "La desensibilización a corto plazo del receptor de angiotensina II de las células glomerulosas suprarrenales de bovinde corresponde a un cambio de un estado de alta a baja afinidad". Endocrinología . 135 (5): 2130–6. doi :10.1210/en.135.5.2130. PMID 7956936.
^ abcdefghijkl Boulpaep EL, Boro WF (2005). Fisiología médica: un enfoque celular y molecular . San Luis, Missouri: Elsevier Saunders. pag. 90.ISBN _1-4160-2328-3.
^ Waltenbaugh C, Doan T, Melvold R, Viselli S (2008). Inmunología . Filadelfia: Wolters Kluwer Health/Lippincott Williams & Wilkins. pag. 20.ISBN _978-0-7817-9543-2.
enlaces externos
Compendio de canales iónicos y base de datos GPCR de IUPHAR Archivado el 23 de marzo de 2019 en Wayback Machine.
Receptomo de membrana plasmática humana Archivado el 15 de septiembre de 2019 en la Wayback Machine.
.svg/1280px-Receptor_(Biochemistry).svg.png)
![{\displaystyle {[{\ce {L}}]+[{\ce {R}}]{\ce {<=>[{K_{d}}]}}[{\text{LR}}]} }](https://wikimedia.org/api/rest_v1/media/math/render/svg/8bc59879baab1d42236bb74c50da94e2ceef5afd)