Una propiedad de una bicapa lipídica es la movilidad relativa (fluidez) de las moléculas lipídicas individuales y cómo esta movilidad cambia con la temperatura. Esta respuesta se conoce como comportamiento de fase de la bicapa. En términos generales, a una temperatura determinada, una bicapa lipídica puede existir en fase líquida o sólida. La fase sólida se denomina comúnmente fase "gel". Todos los lípidos tienen una temperatura característica a la que sufren una transición ( fusión ) de la fase gel a la fase líquida. En ambas fases, las moléculas lipídicas están constreñidas al plano bidimensional de la membrana, pero en las bicapas en fase líquida las moléculas se difunden libremente dentro de este plano. Así, en una bicapa líquida, un lípido determinado intercambiará rápidamente ubicaciones con su vecino millones de veces por segundo y, mediante el proceso de un paseo aleatorio , migrará a largas distancias. [1]
En contraste con esta gran movilidad en el plano, a las moléculas lipídicas les resulta muy difícil cambiar de un lado de la bicapa lipídica al otro. En una bicapa basada en fosfatidilcolina , este proceso normalmente ocurre en un plazo de semanas. [2] Esta discrepancia puede entenderse en términos de la estructura básica de la bicapa. Para que un lípido pase de una valva a otra, su grupo de cabeza hidratado debe cruzar el núcleo hidrofóbico de la bicapa, un proceso energéticamente desfavorable. A diferencia de las bicapas en fase líquida, los lípidos en una bicapa en fase gel están bloqueados en su lugar y no presentan movilidad lateral ni flip-flop. Debido a esta movilidad limitada, las bicapas de gel carecen de una propiedad importante de las bicapas líquidas: la capacidad de volver a sellar pequeños agujeros. Las bicapas en fase líquida pueden curar espontáneamente pequeños huecos, de la misma manera que una película de aceite sobre agua podría fluir para llenar un hueco. Esta funcionalidad es una de las razones por las que las membranas celulares suelen estar compuestas por bicapas en fase fluida. Las limitaciones de movimiento de los lípidos en las bicapas lipídicas también están impuestas por la presencia de proteínas en las membranas biológicas, especialmente en la capa lipídica anular "unida" a la superficie de las proteínas integrales de la membrana .
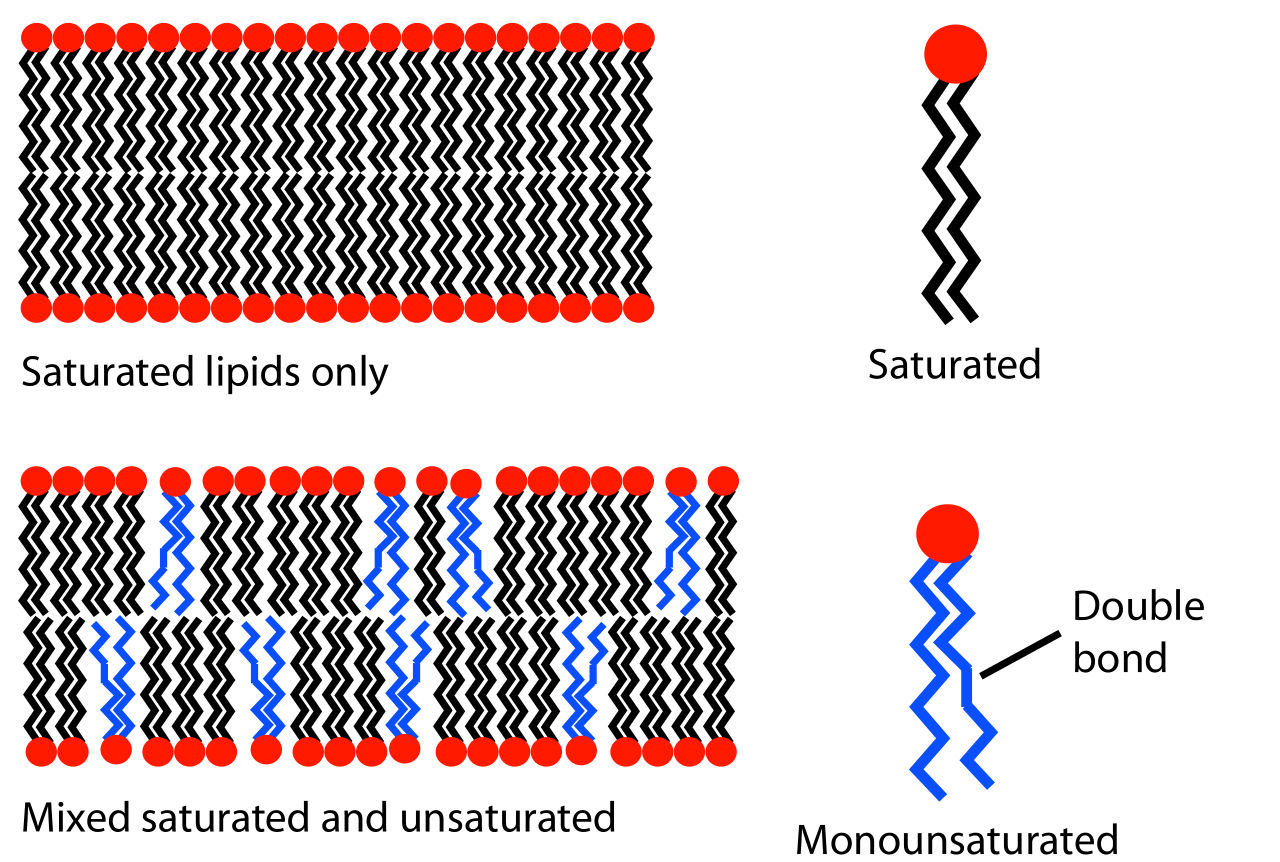
El comportamiento de fase de las bicapas lipídicas está determinado en gran medida por la fuerza de las interacciones atractivas de Van der Waals entre moléculas lipídicas adyacentes. El alcance de esta interacción depende a su vez de la longitud de las colas de lípidos y de su capacidad para agruparse. Los lípidos de cola más larga tienen más área sobre la cual interactuar, aumentando la fuerza de esta interacción y, en consecuencia, disminuyendo la movilidad de los lípidos. Por lo tanto, a una temperatura determinada, un lípido de cola corta será más fluido que un lípido de cola larga idéntico. [3] Otra forma de expresar esto sería decir que la temperatura de transición de la fase gel a líquida aumenta al aumentar el número de carbonos en las cadenas de alcanos lipídicos . Los lípidos de fosfatidilcolina saturados con colas de más de 14 carbonos son sólidos a temperatura ambiente, mientras que aquellos con menos de 14 son líquidos. Este fenómeno es análogo al hecho de que la cera de parafina , que está compuesta de alcanos largos, es sólida a temperatura ambiente, mientras que el octano ( gasolina ), un alcano corto, es líquido.
Además de la longitud de la cadena, la temperatura de transición también puede verse afectada por el grado de insaturación de las colas lipídicas. Un doble enlace insaturado puede producir una torcedura en la cadena del alcano, alterando la estructura periódica regular. Esta interrupción crea un espacio libre adicional dentro de la bicapa que permite una flexibilidad adicional en las cadenas adyacentes. Es esta alteración del empaquetamiento la que conduce a temperaturas de transición más bajas con dobles enlaces crecientes. [3] Este es un efecto particularmente poderoso; Disminuir la longitud total de la cadena en un carbono generalmente altera la temperatura de transición de un lípido en diez grados Celsius o menos, pero agregar un doble enlace simple puede disminuir la temperatura de transición en cincuenta grados o más (ver tabla). Un ejemplo de este efecto se puede observar en la vida cotidiana, ya que la mantequilla , que tiene un gran porcentaje de grasas saturadas , es sólida a temperatura ambiente mientras que el aceite vegetal , que es mayoritariamente insaturado, es líquido.
Las bicapas no necesitan estar compuestas de un solo tipo de lípido y, de hecho, la mayoría de las membranas naturales son una mezcla compleja de diferentes moléculas de lípidos. Estas mezclas a menudo exhiben propiedades intermedias a las de sus componentes, pero también son capaces de producir un fenómeno que no se observa en los sistemas de un solo componente: la separación de fases . Si algunos de los componentes son líquidos a una temperatura determinada mientras que otros están en fase de gel, las dos fases pueden coexistir en poblaciones espacialmente separadas. Esta separación de fases juega un papel crítico en los fenómenos bioquímicos porque los componentes de la membrana, como las proteínas, pueden dividirse en una u otra fase [5] y, por lo tanto, concentrarse o activarse localmente.
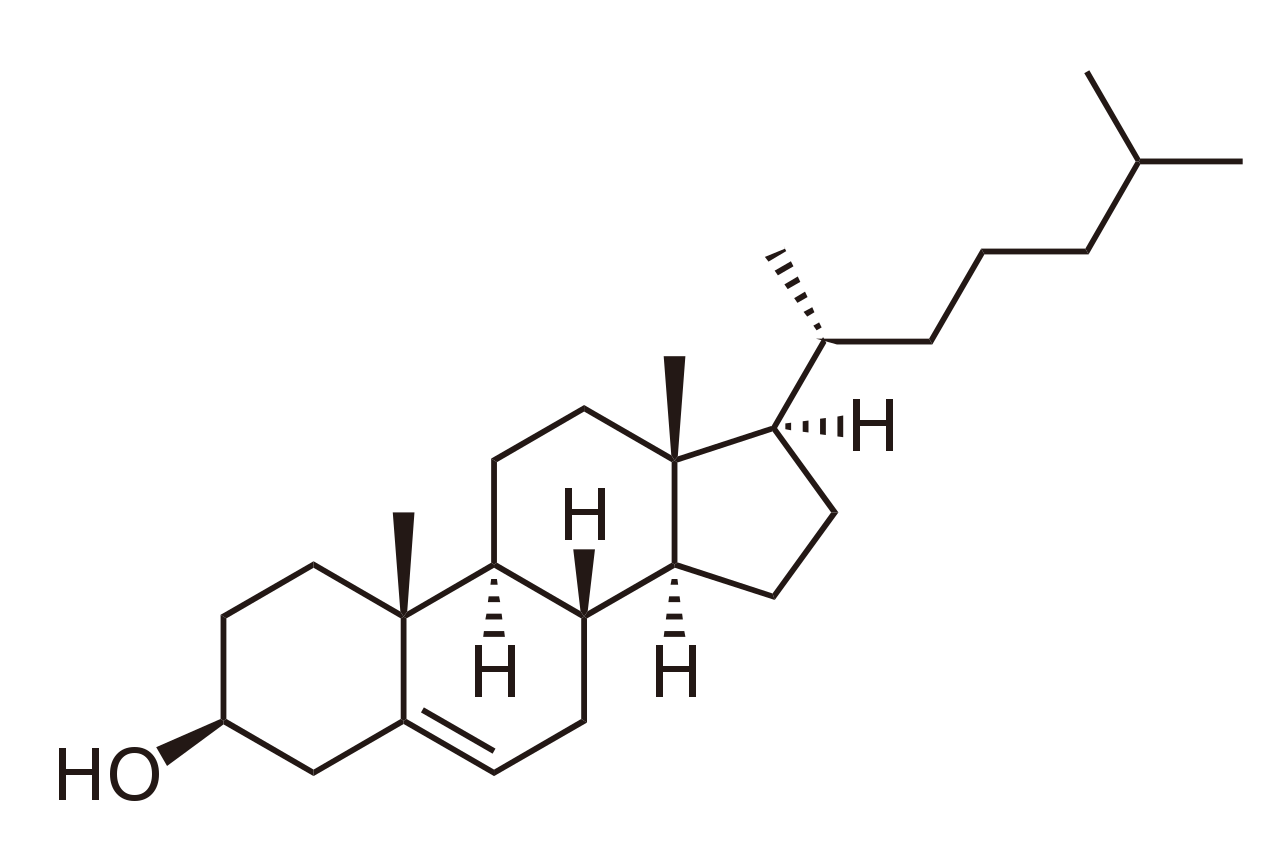
La presencia de colesterol ejerce una influencia profunda pero complicada sobre las propiedades de la bicapa lipídica debido a sus características físicas únicas. Aunque es un lípido, el colesterol guarda poco parecido con un fosfolípido . El dominio hidrófilo del colesterol es bastante pequeño y consta de un solo grupo hidroxilo . Adyacente a este grupo hidroxilo hay una estructura plana rígida compuesta de varios anillos fusionados. En el extremo opuesto de la estructura del anillo hay una cola corta de una sola cadena. Se sabe desde hace décadas que la adición de colesterol a una bicapa en fase fluida disminuye su permeabilidad al agua. [6] [7] Más recientemente se ha demostrado que el modo de esta interacción se debe a que el colesterol se intercala entre las moléculas lipídicas, llenando el espacio libre y disminuyendo la flexibilidad de las cadenas lipídicas circundantes. [8] Esta interacción también aumenta la rigidez mecánica de las bicapas lipídicas de la membrana fluida [9] y disminuye su coeficiente de difusión lateral. [10] Por el contrario, la adición de colesterol a las bicapas en fase de gel altera el orden de empaquetamiento local, aumentando el coeficiente de difusión [10] y disminuyendo el módulo elástico. Las interacciones del colesterol con sistemas de múltiples componentes son aún más complicadas, ya que pueden dar lugar a complejos diagramas de fases . [11] Un sistema de lípidos y colesterol que recientemente se ha estudiado intensamente es la balsa de lípidos. Las balsas lipídicas son dominios de gel enriquecidos con colesterol que han sido potencialmente implicados en ciertos procesos de señalización celular, [12] pero el tema sigue siendo controvertido, y algunos investigadores dudan incluso de su existencia in vivo. [13]
Los liposomas lipídicos mixtos pueden sufrir cambios en diferentes estructuras de dispersión de fases, llamados polimorfismos lipídicos , por ejemplo, micelas esféricas , laminillas de bicapa lipídica y cilindros de fase hexagonal , dependiendo de los cambios físicos y químicos en su microambiente. [14] La temperatura de transición de fase de liposomas y membranas biológicas se puede medir mediante calorimetría , espectroscopia de resonancia magnética y otras técnicas. [15]