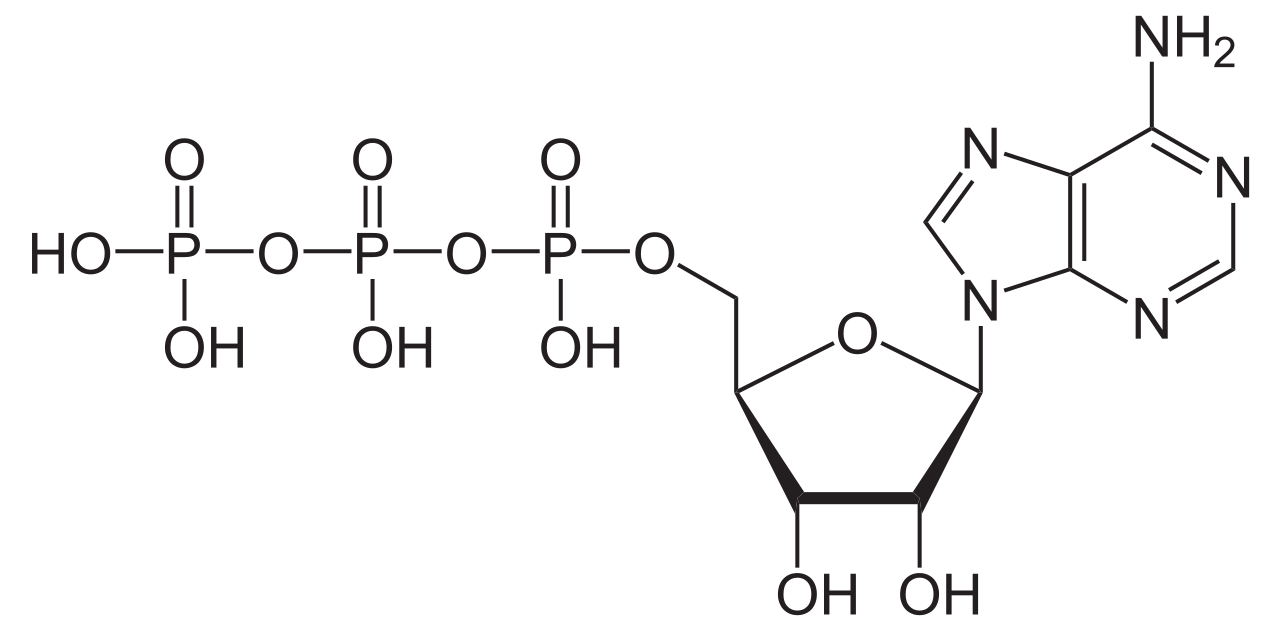

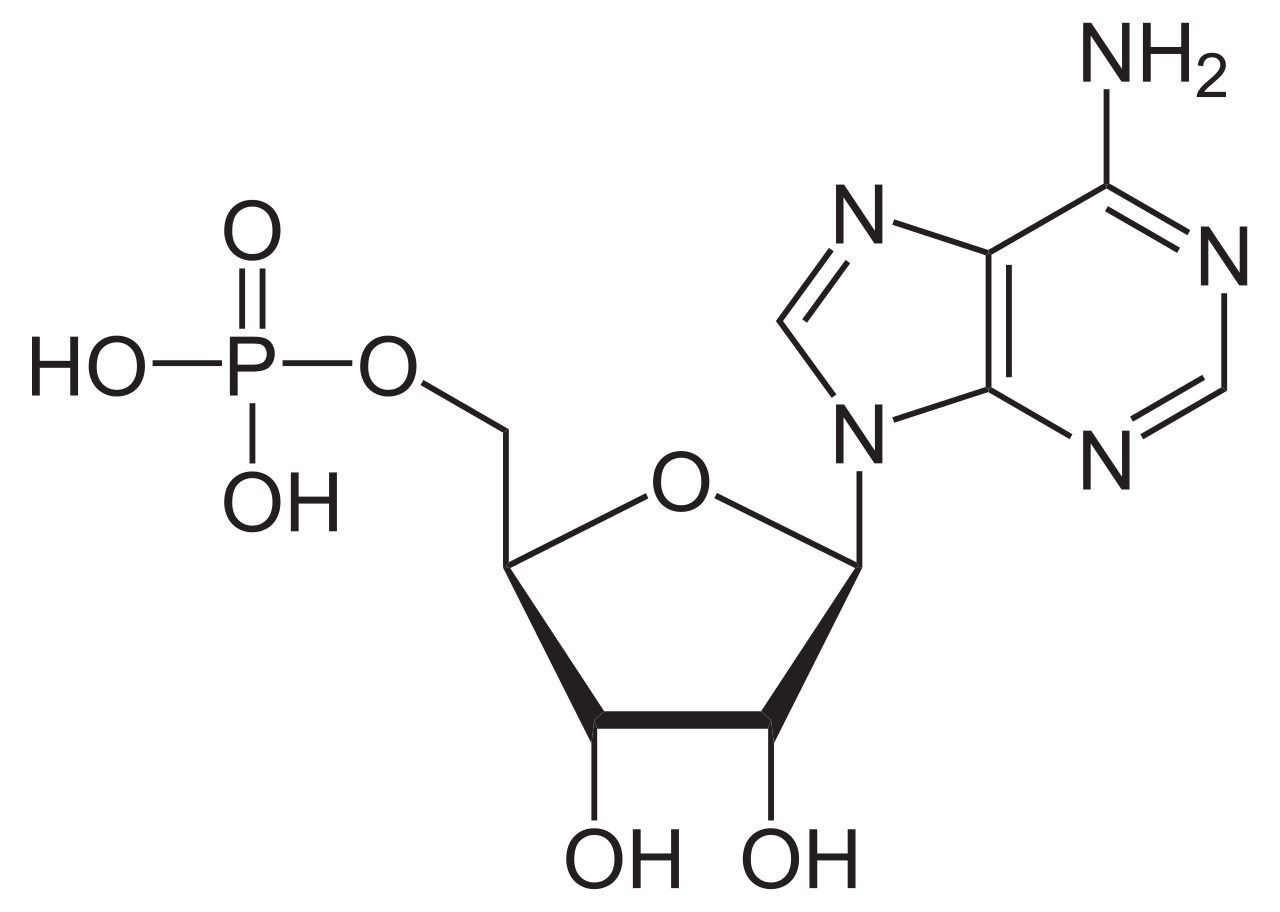
ATPasas ( EC 3.6.1.3, A denosina 5'- Trifosfatasa , adenilpirofosfatasa , ATP monofosfatasa, trifosfatasa, antígeno T SV40, ATP hidrolasa, complejo V (transporte de electrones mitocondriales), ( Ca 2+ + Mg 2+ ) -ATPasa, HCO 3 − -ATPasa, adenosina trifosfatasa) son una clase de enzimas que catalizan la descomposición de ATP en ADP y un ion fosfato libre [1] [2] [3] [4] [5] [6] o la reacción inversa. Esta reacción de desfosforilación libera energía , que la enzima (en la mayoría de los casos) aprovecha para impulsar otras reacciones químicas que de otro modo no ocurrirían. Este proceso es ampliamente utilizado en todas las formas de vida conocidas .
Algunas de estas enzimas son proteínas de membrana integrales (ancladas dentro de membranas biológicas ) y mueven solutos a través de la membrana, generalmente en contra de su gradiente de concentración. Se denominan ATPasas transmembrana.

Las ATPasas transmembrana importan metabolitos necesarios para el metabolismo celular y exportan toxinas, desechos y solutos que pueden obstaculizar los procesos celulares. Un ejemplo importante es la bomba de sodio-potasio (Na + /K + ATPasa) que mantiene el potencial de membrana celular . Otro ejemplo es la hidrogenopotasio ATPasa (H + /K + ATPasa o bomba de protones gástrica) que acidifica el contenido del estómago. La ATPasa está genéticamente conservada en los animales; por lo tanto, los cardenólidos , que son esteroides tóxicos producidos por plantas que actúan sobre las ATPasas, producen toxinas animales generales y efectivas que actúan de manera dependiente de la dosis. [7]
Además de los intercambiadores, otras categorías de ATPasa transmembrana incluyen cotransportadores y bombas (sin embargo, algunos intercambiadores también son bombas). Algunos de ellos, como la Na + /K + ATPasa, provocan un flujo neto de carga, pero otros no. Estos se denominan transportadores electrogénicos y transportadores electroneutros, respectivamente. [8]
"La adenosina trifosfatasa transportadora de cobre unida a la membrana (Cu-ATPasa), que une selectivamente iones de cobre, transporta iones de cobre dentro y fuera de las células (Harris et al. 1998)". Fuente: https://www.atsdr.cdc.gov/ToxProfiles/tp132.pdf pág. 73
Los motivos de Walker son un motivo de secuencia de proteínas revelador de la unión e hidrólisis de nucleótidos. Más allá de esta amplia función, los motivos de Walker se pueden encontrar en casi todas las ATPasas naturales, con la notable excepción de las tirosina quinasas . [9] Los motivos de Walker comúnmente forman una hoja Beta -convertida en hélice Alfa- que se autoorganiza como un Nido (motivo estructural proteico) . Se cree que esto se debe a que las ATPasas modernas evolucionaron a partir de pequeños péptidos de unión a NTP que tenían que autoorganizarse. [10]
El diseño de proteínas ha podido replicar la función de la ATPasa (débilmente) sin utilizar secuencias o estructuras de ATPasa naturales. Es importante destacar que, si bien todas las ATPasas naturales tienen alguna estructura de hoja beta, la "ATPasa alternativa" diseñada carece de estructura de hoja beta, lo que demuestra que esta función esencial para la vida es posible con secuencias y estructuras que no se encuentran en la naturaleza. [11]
La ATPasa (también llamada F 0 F 1 -ATP Sintasa) es un complejo de transferencia de carga que cataliza el ATP para realizar la síntesis de ATP moviendo iones a través de la membrana. [12]
El acoplamiento de la hidrólisis y el transporte de ATP es una reacción química en la que se transporta un número fijo de moléculas de soluto por cada molécula de ATP hidrolizada; para el intercambiador Na + /K + , son tres iones Na + fuera de la célula y dos iones K+ dentro por cada molécula de ATP hidrolizada.
Las ATPasas transmembrana utilizan la energía potencial química del ATP realizando trabajo mecánico: transportan solutos en la dirección opuesta a su dirección de movimiento termodinámicamente preferida, es decir, desde el lado de la membrana con baja concentración hacia el lado con alta concentración. Este proceso se conoce como transporte activo .
Por ejemplo, la inhibición de las H + -ATPasas vesiculares daría como resultado un aumento del pH dentro de las vesículas y una caída del pH del citoplasma.
Todas las ATPasas comparten una estructura básica común. Cada ATPasa rotatoria se compone de dos componentes principales: F 0 /A 0 /V 0 y F 1 /A 1 /V 1 . Están conectados por 1 a 3 tallos para mantener la estabilidad, controlar la rotación y evitar que giren en la otra dirección. Se utiliza un tallo para transmitir el par. [13] El número de tallos periféricos depende del tipo de ATPasa: las F-ATPasas tienen uno, las A-ATPasas tienen dos y las V-ATPasas tienen tres. El dominio catalítico F 1 está ubicado en el lado N de la membrana y participa en la síntesis y degradación de ATP y participa en la fosforilación oxidativa . El dominio transmembrana F 0 participa en el movimiento de iones a través de la membrana. [12]
La F 0 F 1 -ATPasa bacteriana consta del dominio F 1 soluble y el dominio F 0 transmembrana , que se compone de varias subunidades con estequiometría variable. Hay dos subunidades, γ y ε, que forman el tallo central y están vinculadas a F 0 . F 0 contiene un oligómero de la subunidad c en forma de anillo (anillo c). La subunidad α está cerca de la subunidad b 2 y forma el tallo que conecta las subunidades transmembrana con las subunidades α3β3 y δ. Las F-ATP sintasas son idénticas en apariencia y función, excepto la F 0 F 1 -ATP sintasa mitocondrial, que contiene de 7 a 9 subunidades adicionales. [12]
El potencial electroquímico es lo que hace que el anillo C gire en el sentido de las agujas del reloj para la síntesis de ATP. Esto hace que el tallo central y el dominio catalítico cambien de forma. Al girar el anillo C se producen tres moléculas de ATP, lo que luego hace que H + se mueva del lado P de la membrana al lado N de la membrana. La rotación en sentido antihorario del anillo C es impulsada por la hidrólisis del ATP y los iones se mueven del lado N al lado P, lo que ayuda a generar potencial electroquímico. [12]
La ATP sintasa de mitocondrias y cloroplastos es una enzima anabólica que aprovecha la energía de un gradiente de protones transmembrana como fuente de energía para agregar un grupo fosfato inorgánico a una molécula de difosfato de adenosina (ADP) para formar una molécula de trifosfato de adenosina (ATP).
Esta enzima funciona cuando un protón desciende en el gradiente de concentración, lo que le da a la enzima un movimiento giratorio. Este movimiento giratorio único une ADP y P para crear ATP.
La ATP sintasa también puede funcionar a la inversa, es decir, utilizar la energía liberada por la hidrólisis del ATP para bombear protones contra su gradiente electroquímico.
Hay diferentes tipos de ATPasas, que pueden diferir en función (síntesis y/o hidrólisis de ATP), estructura (las ATPasas F, V y A contienen motores rotativos) y en el tipo de iones que transportan.
Las P-ATPasas (a veces conocidas como ATPasas E1-E2) se encuentran en bacterias y también en membranas plasmáticas y orgánulos de eucariotas. Su nombre se debe a la unión breve del fosfato inorgánico a los residuos de aspartato en el momento de la activación. La función de la P-ATPasa es transportar una variedad de compuestos diferentes, como iones y fosfolípidos, a través de una membrana utilizando la hidrólisis de ATP para obtener energía. Hay muchas clases diferentes de P-ATPasas, que transportan un tipo específico de ion. Las P-ATPasas pueden estar compuestas por uno o dos polipéptidos y normalmente pueden adoptar dos conformaciones principales, E1 y E2.
{kind=link}