Panderichthys es un género de sarcopterigio (pez con aletas lobuladas) extinto del período Devónico tardío , hace unos 380 millones de años . Panderichthys , que fue recuperado de depósitos del Frasniano (principios del Devónico tardío) en Letonia , está representado por dos especies. P. stolbovi se conoce solo a partir de algunos fragmentos de hocico y una mandíbula inferior incompleta. P. rhombolepis se conoce a partir de varios especímenes más completos. Aunque probablemente pertenece a un grupo hermano de los primeros tetrápodos , Panderichthys exhibe una gama de características transicionales entre los peces tristicópteridos de aletas lobuladas (por ejemplo, Eusthenopteron ) y los primeros tetrápodos. [1] Recibe su nombre en honor al paleontólogo alemán-báltico Christian Heinrich Pander .En 2010 se informaronposibles huellas de tetrápodos que datan de antes de la aparición de Panderichthys en el registro fósil, lo que sugiere que Panderichthys no es un ancestro directo de los tetrápodos, pero no obstante muestra los rasgos que evolucionaron durante la evolución pez-tetrápodo [2].
El Panderichthys está representado por dos especies diferentes: Panderichthys rhombolepis y Panderichthys stobolvi . P. rhombolepis fue descubierto por Gross en 1930 y P. stobolvi fue descubierto y descifrado por Emilia Vorobyeva en 1960. P. rhombolepis fue descubierto en Lode, Letonia, dentro de los depósitos frasnianos y, según PE Ahlberg, definitivamente se puede encontrar en otros depósitos frasnianos en Letonia. Aunque los fósiles de Panderichthys se conocen desde hace mucho tiempo, solo recientemente se han examinado en profundidad. [3] La primera vez que se reconoció que estaban filogenéticamente más cerca de los tetrápodos que de los peces fue por Shultze y Arsenault en 1985.

Panderichthys es un pez de 1,5 a 2 m (4 pies 11 pulgadas - 6 pies 7 pulgadas) de largo [4] con una gran cabeza similar a la de un tetrápodo que es aplanada, estrecha en el hocico y ancha en la espalda. La articulación intracraneal, que es característica de la mayoría de los peces de aletas lobuladas, se ha perdido de los elementos externos del cráneo, pero todavía está presente en la caja craneana. Los patrones de los huesos externos en el techo del cráneo y las mejillas son más similares a los de los tetrápodos primitivos que a los de otros peces de aletas lobuladas. [1] Las cualidades transicionales de Panderichthys también son evidentes en el resto del cuerpo. Carece de las aletas dorsal y anal ( aleta de pez ) y su cola es más parecida a la de los tetrápodos primitivos que a las aletas caudales de otros peces de aletas lobuladas. Los hombros exhiben varias características similares a las de los tetrápodos, mientras que el húmero es más largo que los que se encuentran en otros peces de aletas lobuladas. La columna vertebral está osificada en toda su longitud y las vértebras son comparables a las de los primeros tetrápodos. [1] Por otra parte, las partes distales de las aletas delanteras son diferentes a las de los tetrápodos. Como sería de esperar de una aleta, hay numerosos lepidotriquios (radios de la aleta largos y delgados).
Panderichthys posee muchas características que pueden considerarse una forma intermedia durante la evolución de pez a tetrápodo y muestra algunas características que son más derivadas de lo que indica su posición filogenética, mientras que otras son más basales. La forma corporal de Panderichthys y Tiktaalik representa un paso importante en la transición de pez a tetrápodo e incluso pudieron desplazarse en tierra. [5] Según Shultz y Trueb, Panderichthys comparte diez características con los tetrápodos: [6]
Una de las características transicionales clave de Panderichthys es su húmero . Durante la transición de pez a tetrápodo, las extremidades comenzaron a moverse y se ubicaron en un ángulo recto con el cuerpo en lugar de estar orientadas hacia el extremo posterior. Como resultado, los músculos se volvieron perpendiculares al cuerpo y causaron que las extremidades se movieran en un patrón más anteroposterior y dorsoventral. Esto a su vez afectó la forma del húmero y, como resultado, los primeros tetrápodos tienen un húmero en forma de L. [7] Debido a un descubrimiento reciente de un húmero de Panderichthys que no estaba aplanado, el espécimen pudo analizarse con mucho más detalle. El húmero de Panderichthys muestra una variedad de características que incluyen algunas que son tanto primitivas como derivadas. A pesar de estar ubicado como basal a Tiktaalik , el húmero de Panderichthys tiene características que son más derivadas, pero en general es muy similar. Tanto Panderichthys como Tiktaalik tienen húmeros aplanados dorsoventralmente con un entepicóndilo en forma de cuchilla que se curva ventralmente, facetas epipodiales separadas, un proceso del dorsal ancho y un proceso ectepicondíleo que es paralelo al margen preaxial. Los húmeros de ambas especies se consideran formas de transición porque tienen forma casi de L, un proceso del dorsal ancho bajo, un entepicóndilo bajo y un canal entepicondilar intermedio. El húmero de Panderichthys es más derivado que el de Tiktaalik debido a la presencia de una faceta radial orientada más preaxialmente, así como un eje más delgado. Una característica que es única de Panderichthys es que el entepicóndilo no se proyecta tanto como las facetas epipodiales y la cresta humeral no se adentra en el entepicóndilo. [8] El resultado del análisis del húmero de Panderichthys es que la transición del húmero de los organismos similares a peces a los tetrápodos ocurrió mucho más lentamente de lo que se pensaba anteriormente y Panderichthys ahora proporciona una base para determinar muchas autapomorfías. [8] Debido a la orientación de la aleta hacia el extremo posterior, la actitud de la extremidad es más horizontal que vertical y el espacio operativo en el que actúa está al nivel de la articulación del hombro, lo que hace que los músculos tiren en ángulo recto con el cuerpo. Esto resultó en la capacidad de Panderichthys de sostener su gran cabeza, probablemente para respirar. [9]

Otra característica clave de Panderichthys es su forma intermedia durante la evolución de los dígitos. En el pasado se creía que los dígitos y los dedos no tenían una parte análoga en los peces sarcopterigios y eran novedades evolutivas. Sin embargo, un nuevo examen [10] de los fósiles existentes de Panderichthys utilizando un escáner de TC muestra al menos cuatro huesos radiales distales muy claramente diferenciados al final de la estructura esquelética de la aleta. Este estudio, realizado por Boisvert et al. en 2008, examinó las aletas pectorales de Panderichthys y descubrió que las aletas de Panderichthys están orientadas anteroposteriormente, lo que es diferente de las extremidades de los tetrápodos que se proyectan en un ángulo desde el cuerpo. El húmero, el radio y el cúbito son todos reconocibles como análogos a las partes de los tetrápodos. [11]
Las tomografías computarizadas permitieron a los autores ver debajo de las escamas y lepidotrichia (radios de la aleta), descubriendo por primera vez el endoesqueleto de la aleta distal. La tomografía computarizada mostró un cúbito , un carpo (hueso de la muñeca) en forma de bloque que se articula con el cúbito y dos radiales terminales. La muñeca también incluía un intermedio más delgado , que se articula en línea con la cresta lateral del cúbito. La tomografía computarizada descubrió además radiales que pueden interpretarse como dígitos, lo que cuestiona la hipótesis de que los dígitos son estructuras completamente nuevas en los tetrápodos. Estos huesos similares a dedos no muestran desarrollo muscular ni articulaciones y son extremadamente pequeños, pero no obstante muestran una forma intermedia entre las aletas completamente parecidas a las de los peces y los tetrápodos. Similar al húmero, Panderichthys también tiene una característica más derivada similar a los tetrápodos y a diferencia de Tiktaalik : el cúbito es significativamente más largo que el cúbito. [11]

La cintura pélvica (cadera) y las aletas pélvicas de Panderichthys representan un intermedio en la evolución pez-tetrápodo. Durante la evolución pez-tetrápodo la cintura pélvica se convirtió en una estructura de soporte de peso cuando se desarrollaron el íleon, el contacto mesoventral de los lados de la cintura, un íleon y una costilla sacra. El fémur y el húmero se hicieron más largos y el radio/cúbito y la tibia/peroné se volvieron más iguales en longitud. En general, la cintura pélvica en Panderichthys es más primitiva que la cintura escapular. Esto se debe a que el húmero de Panderichthys tiene una forma que es más intermedia, mientras que el fémur es más primitivo debido a la relación de longitud con el peroné y a que carece de una lámina aductora y cresta. Esto implica que Panderichthys no era capaz de moverse con sus extremidades traseras como los tetrápodos debido a sus pequeñas aletas pélvicas, cintura pélvica que no soporta peso, acetebelo orientado posteriormente y flexión limitada de rodillas y codos. [12] Boisvert describe la locomoción de Panderichthys como posiblemente usando una de sus aletas pectorales para anclarse mientras la ondulación de lado a lado impulsa el cuerpo hacia adelante. [12]
La caja craneal de Panderichthys muestra un intermedio clave dentro de la secuencia de evolución de los peces. Desde el exterior, Panderichthys tiene una cabeza similar a la de un tetrápodo, pero en realidad conserva una articulación intracraneal que es característica de los peces. Panderichthys comparte muchas características con el osteolepiforme Eusthenopteron , como procesos hiomandibulares y basipterigoideos similares. Aunque su cabeza tiene una forma similar a la de un tetrápodo, los cráneos de los tetrápodos carecen de comisura lateral, surco yugular, fenestra basicraneal, placa arqueada y articulación intracraneal, todos los cuales están presentes en Panderichthys . Lo que esto significa es que no hubo un cambio importante en la construcción de la caja craneal desde el primer sarcopterigio, sino que solo hubo cambios en la forma del cráneo. Esto implica que la evolución de la caja craneal durante la transición de pez a tetrápodo fue muy rápida y parece mostrar el mismo ritmo que la evolución de la cintura pélvica. En general, Panderichthys demuestra que la estructura de la caja craneana evolucionó mucho más lentamente que la morfología externa del cráneo que creó la apariencia tetrápoda de la cabeza. [13]
En cuanto a la mandíbula inferior y la dentición, la mandíbula inferior es similar a la de los ripidistios y está compuesta por un dentario con dientes, cuatro intradentarios, un prearticular lingual, tres coronoides y una placa adsinfisaria dorsalmente. Además, los dientes son de estructura poliplocodonte. [6]
Como intermediario en la evolución pez-tetrápodo, Panderichthys tenía la capacidad de respirar aire. La tendencia desde los primeros sarcopterigios hasta los primeros tetrápodos fue un aumento en el tamaño de la cámara espiracular y su apertura al exterior. En comparación con Eusthenopteron , la cámara espiracular de Panderichthys está muy expandida y los hiomandibulares son más cortos en comparación con los de los peces. La serie opercular también era más corta en comparación con otros osteolepiformes. [3] Panderichthys también tiene una única abertura nasal externa y una coana palatina. A diferencia de los osteolepiformes anteriores, la coana palatina es alargada y la lámina nariocoanal es estrecha. Junto con la cámara espiracular, esta característica en Panderichthys puede considerarse transicional durante la evolución de pez a tetrápodos. [6] Los sarcopterigios como Panderichthys pueden considerarse al menos respiradores de aire facultativos y demuestran una forma intermedia a medida que la respiración aérea se hacía más abundante. [3]

En enero de 2010, Nature informó sobre huellas de tetrápodos bien conservadas y "con datación segura" de sedimentos de llanuras de marea marinas polacas de aproximadamente 397 millones de años de antigüedad. [2] Estas huellas fósiles sugieren que un grupo de tetrápodos de dos metros de largo vivió en áreas intermareales o lagunares completamente marinas en la costa sur de Laurussia durante la época en que vivieron los elpistostegidos. Esto implica que Panderichthys no es un fósil de transición y representa su propia morfología adaptativa. Por lo tanto, Panderichthys solo puede ser una "reliquia de supervivencia tardía", [14] que muestra rasgos que evolucionaron durante la transición de criaturas similares a peces a tetrápodos, pero cuya fecha no refleja esa transición. Las huellas "obligan a una reevaluación radical del momento, la ecología y el entorno ambiental de la transición pez-tetrápodo, así como de la integridad del registro fósil corporal". [2]
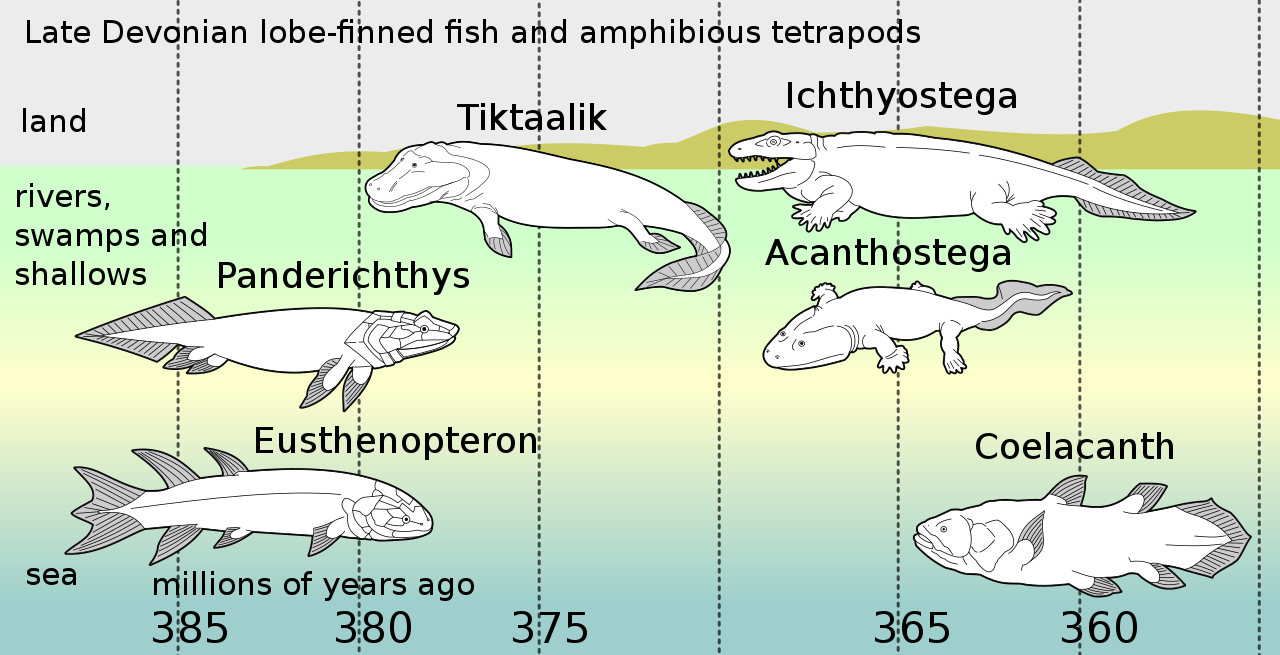
Panderichthys estuvo vivo durante el Devónico tardío ( Frasniano ) en Lode, Letonia. Se sabe que Lode es un entorno marino marginal y se ha planteado la hipótesis de que Panderichthys estaba adaptado para el movimiento en aguas poco profundas y llenas de escombros. [3] Panderichthys fue recolectado en depósitos que anteriormente se creía que eran de una cuenca de agua dulce tranquila, pero que se ha demostrado que son de llanuras de marea poco profundas o un estuario. La Formación Lode, donde se encontró P. rhombolepis , se encuentra dentro de una capa de 200 metros de espesor compuesta de arenisca de grano fino y arcilla junto con arcillas finamente dispersas. Casi todos los taxones principales de vertebrados del Devónico tardío están representados dentro de la Formación Lode. La mayoría de los especímenes están bien conservados debido a las condiciones anaeróbicas del sustrato, así como al entierro rápido en depresiones en las laderas del delta submarino. P. rhombolepis fue descubierto en la formación regional Gauja dentro de la sección inferior del Frasniano. La tafocoenosis se caracterizó por estar en arcilla finamente desplazada y arcilla limosa, así como por una baja actividad de agua. Dentro de este ambiente se ha planteado la hipótesis de que P. rhombolepis era un gran depredador y se alimentaba de dípteros, sarcopterigios pequeños y juveniles y Latvius . [15] Los vertebrados asociados encontrados en los mismos depósitos incluyen un pez sin mandíbula acorazado ( Psammolepis ), dos placodermos ( Asterolepis y Plourdosteus ), un acantódido no identificado, un aleta lobulada porolepiforme ( Laccognathus ), un pez pulmonado ( Dipterus ) y otro elpistostegaliano ( Livoniana ). [1]

Durante el Frasniano, en el que Panderichthys estaba vivo, hubo una caída del oxígeno en la atmósfera, así como un aumento en la abundancia de plantas. Debido a que el oxígeno es mucho menos soluble en agua que el aire, la disminución del oxígeno en la atmósfera habría causado que las concentraciones de oxígeno en cualquier tipo de agua disminuyeran sustancialmente. Esto, a su vez, habría hecho que cualquier animal acuático que pudiera respirar aire tuviera una ventaja y fuera más probable que prosperara. [3] Además de su capacidad para moverse en aguas poco profundas, Panderichthys también podía respirar aire. Sus fuertes aletas pectorales, en teoría, podrían permitirle sostener su cabeza en aguas poco profundas y respirar. El agrandamiento de la cámara espiracular en sí, así como su apertura al exterior, sugieren que Panderichthys fue parte de una transición hacia una mayor capacidad para respirar aire que se completó en los tetrápodos. [3]