La evolución de los tetrápodos comenzó hace unos 400 millones de años en el Período Devónico , y los primeros tetrápodos evolucionaron a partir de peces con aletas lobuladas . [1] Los tetrápodos (según la definición basada en la apomorfía utilizada en esta página) se clasifican como animales de la superclase biológica Tetrapoda , que incluye a todos los anfibios , reptiles , aves y mamíferos vivos y extintos . Si bien la mayoría de las especies actuales son terrestres, hay poca evidencia que respalde la idea de que alguno de los primeros tetrápodos pudiera moverse sobre la tierra, ya que sus extremidades no podrían haber mantenido sus secciones medias separadas del suelo y las huellas conocidas no indican que arrastraran sus vientres. Presumiblemente, las huellas fueron hechas por animales que caminaban por el fondo de cuerpos de agua poco profundos. [2] Los ancestros acuáticos específicos de los tetrápodos y el proceso por el cual ocurrió la colonización terrestre siguen sin estar claros. Son áreas de investigación activa y debate entre los paleontólogos en la actualidad.
La mayoría de los anfibios actuales siguen siendo semiacuáticos, viviendo la primera etapa de sus vidas como renacuajos similares a los peces . Varios grupos de tetrápodos, como las serpientes y los cetáceos , han perdido algunas o todas sus extremidades. Además, muchos tetrápodos han regresado a vidas parcialmente o completamente acuáticas a lo largo de la historia del grupo (los ejemplos modernos de tetrápodos completamente acuáticos incluyen a los cetáceos y los sirenios ). Los primeros retornos a un estilo de vida acuático pueden haber ocurrido ya en el Período Carbonífero [3], mientras que otros retornos ocurrieron tan recientemente como el Cenozoico , como en el caso de los cetáceos, los pinnípedos [4] y varios anfibios modernos [5] .
El cambio de un plan corporal para respirar y navegar en el agua a un plan corporal que permite al animal moverse en la tierra es uno de los cambios evolutivos más profundos conocidos. [6] También es uno de los mejor comprendidos, en gran parte gracias a una serie de importantes hallazgos de fósiles de transición a finales del siglo XX combinados con un mejor análisis filogenético. [1]
El período Devónico se conoce tradicionalmente como la "Era de los Peces", ya que marca la diversificación de numerosos grupos de peces importantes, tanto actuales como extintos. [7] Entre ellos se encontraban los primeros peces óseos , que se diversificaron y se extendieron en ambientes de agua dulce y salobre al comienzo del período. Los primeros tipos se parecían a sus ancestros cartilaginosos en muchas características de su anatomía, incluida una aleta caudal similar a la de un tiburón, intestino en espiral, grandes aletas pectorales endurecidas en el frente por elementos esqueléticos y un esqueleto axial en gran parte no osificado . [8]
Sin embargo, tenían ciertos rasgos que los separaban de los peces cartilaginosos, rasgos que se volverían fundamentales en la evolución de las formas terrestres. Con la excepción de un par de espiráculos , las branquias no se abrían individualmente al exterior como lo hacen en los tiburones; más bien, estaban encerradas en una cámara branquial reforzada por huesos membranosos y cubierta por un opérculo óseo , con una sola abertura al exterior. El hueso cleithrum , que forma el margen posterior de la cámara branquial, también funcionaba como anclaje para las aletas pectorales. Los peces cartilaginosos no tienen tal anclaje para las aletas pectorales. Esto permitió una articulación móvil en la base de las aletas en los primeros peces óseos, y más tarde funcionaría en una estructura de soporte de peso en los tetrápodos. Como parte de la armadura general de escamas romboides de la cósmina , el cráneo tenía una cubierta completa de hueso dérmico , que constituía un techo craneal sobre el cráneo interior cartilaginoso, que por lo demás era similar al de un tiburón . Es importante destacar que también tenían un par de pulmones ventrales pareados , [9] una característica de la que carecen los tiburones y las rayas.
Se suponía que los peces evolucionaron en gran medida alrededor de los arrecifes , pero desde su origen hace unos 480 millones de años, vivían en entornos cercanos a la costa como áreas intermareales o lagunas permanentemente poco profundas y no comenzaron a proliferar en otros biotopos hasta 60 millones de años después. Unos pocos se adaptaron a aguas más profundas, mientras que las formas sólidas y de constitución pesada se quedaron donde estaban o migraron al agua dulce. [10] [11] El aumento de la productividad primaria en la tierra durante el Devónico tardío cambió los ecosistemas de agua dulce. Cuando los nutrientes de las plantas se liberaron en lagos y ríos, fueron absorbidos por microorganismos que a su vez fueron comidos por invertebrados, que sirvieron como alimento para los vertebrados. Algunos peces también se volvieron detritívoros . [12] Los primeros tetrápodos desarrollaron una tolerancia a entornos que variaban en salinidad, como estuarios o deltas. [13]
El pulmón/ vejiga natatoria se originó como una extensión del intestino, formando una vejiga llena de gas sobre el sistema digestivo. En su forma primitiva, la vejiga natatoria estaba abierta al tubo digestivo , una condición llamada fisostoma y todavía se encuentra en muchos peces. [14] La función principal de la vejiga natatoria no es del todo segura. Una consideración es la flotabilidad . La pesada armadura de escamas de los primeros peces óseos ciertamente pesaría a los animales. En los peces cartilaginosos, que carecen de vejiga natatoria, los tiburones de mar abierto necesitan nadar constantemente para evitar hundirse en las profundidades, y las aletas pectorales proporcionan elevación . [15] Otro factor es el consumo de oxígeno. El oxígeno ambiental era relativamente bajo en el Devónico temprano, posiblemente alrededor de la mitad de los valores modernos. [16] Por unidad de volumen, hay mucho más oxígeno en el aire que en el agua, y los vertebrados (especialmente los nectónicos ) son animales activos con un mayor requerimiento de energía en comparación con los invertebrados de tamaños similares. [17] [18] El Devónico vio niveles crecientes de oxígeno que abrieron nuevos nichos ecológicos al permitir que los grupos capaces de explotar el oxígeno adicional se desarrollaran en animales activos y de gran tamaño. [16] Particularmente en hábitats de pantanos tropicales, el oxígeno atmosférico es mucho más estable y puede haber impulsado una dependencia de proto-pulmones (que realizan esencialmente un tipo evolucionado de respiración enteral ) en lugar de branquias para la absorción primaria de oxígeno. [19] [20] Al final, tanto la flotabilidad como la respiración pueden haber sido importantes, y algunos peces fisóstomos modernos de hecho usan sus vejigas para ambas.
Para funcionar en el intercambio de gases, los pulmones necesitan un suministro de sangre. En los peces cartilaginosos y teleósteos , el corazón se encuentra bajo en el cuerpo y bombea sangre hacia adelante a través de la aorta ventral , que se divide en una serie de arcos aórticos pareados, cada uno correspondiente a un arco branquial . [21] Los arcos aórticos luego se fusionan por encima de las branquias para formar una aorta dorsal que suministra al cuerpo sangre oxigenada. En los peces pulmonados , los bowfin y los bichires , la vejiga natatoria es irrigada con sangre por arterias pulmonares pareadas que se ramifican desde el arco aórtico posterior (sexto). [22] El mismo patrón básico se encuentra en el pez pulmonado Protopterus y en las salamandras terrestres , y probablemente fue el patrón encontrado en los ancestros inmediatos de los tetrápodos, así como en los primeros tetrápodos. [23] En la mayoría de los demás peces óseos, la vejiga natatoria es irrigada con sangre por la aorta dorsal. [22]
Para que los pulmones permitan el intercambio de gases, primero deben tener gas en ellos. En los tetrápodos modernos, se conservan tres mecanismos respiratorios importantes de sus ancestros tempranos, el primero es un sistema de detección de CO 2 /H+. En la respiración de los tetrápodos modernos, el impulso de respirar se desencadena por una acumulación de CO 2 en el torrente sanguíneo y no por una falta de O 2 . [24] Un sistema de detección de CO 2 /H+ similar se encuentra en todos los osteíctios , lo que implica que el último ancestro común de todos los osteíctios necesitaba este tipo de sistema de detección. [24] [25] El segundo mecanismo para respirar es un sistema surfactante en los pulmones para facilitar el intercambio de gases. Esto también se encuentra en todos los osteíctios, incluso en aquellos que son casi completamente acuáticos. [26] [27] La naturaleza altamente conservada de este sistema sugiere que incluso los osteíctios acuáticos tienen cierta necesidad de un sistema surfactante, lo que puede parecer extraño ya que no hay gas bajo el agua. El tercer mecanismo de la respiración es el movimiento real de la respiración. Este mecanismo es anterior al último ancestro común de los osteíctios, como se puede observar en Lampetra camtshatica , el clado hermano de los osteíctios. En las lampreas, este mecanismo toma la forma de una "tos", donde la lamprea sacude su cuerpo para permitir que el agua fluya a través de sus branquias. Cuando los niveles de CO2 en la sangre de la lamprea suben demasiado, se envía una señal a un generador de patrones central que hace que la lamprea "tosa" y permita que el CO2 salga de su cuerpo. [28] [29] Este vínculo entre el sistema de detección de CO2 y el generador de patrones central es extremadamente similar al vínculo entre estos dos sistemas en los tetrápodos, lo que implica homología.
Las fosas nasales de la mayoría de los peces óseos difieren de las de los tetrápodos. Normalmente, los peces óseos tienen cuatro fosas nasales (aberturas nasales), una naris detrás de la otra en cada lado. A medida que el pez nada, el agua fluye hacia el par delantero, a través del tejido olfativo y hacia afuera a través de las aberturas posteriores. Esto es cierto no solo para los peces con aletas radiadas, sino también para el celacanto , un pez incluido en los sarcopterigios , el grupo que también incluye a los tetrápodos. En contraste, los tetrápodos tienen solo un par de narinas externas, pero también lucen un par de narinas internas, llamadas coanas , que les permiten tomar aire a través de la nariz. Los peces pulmonados también son sarcopterigios con fosas nasales internas, pero estas son lo suficientemente diferentes de las coanas de los tetrápodos como para que desde hace mucho tiempo se las haya reconocido como un desarrollo independiente. [30]
La evolución de las fosas nasales internas de los tetrápodos fue objeto de acalorados debates en el siglo XX. Las fosas nasales internas podrían ser un conjunto de las externas (que se suele suponer que son el par posterior) que han migrado a la boca, o el par interno podría ser una estructura recién evolucionada. Sin embargo, para dar paso a una migración, los dos huesos que llevan los dientes de la mandíbula superior, el maxilar y el premaxilar , tendrían que separarse para dejar pasar la fosa nasal y luego volver a unirse; hasta hace poco, no había evidencia de una etapa de transición, con los dos huesos desconectados. Dicha evidencia está ahora disponible: un pequeño pez de aletas lobuladas llamado Kenichthys , encontrado en China y datado en alrededor de 395 millones de años, representa la evolución "atrapada en medio del acto", con el maxilar y el premaxilar separados y una abertura -la incipiente coana- en el labio entre los dos huesos. [31] Kenichthys está más estrechamente relacionado con los tetrápodos que el celacanto, [32] que solo tiene fosas nasales externas; Representa, por tanto, una etapa intermedia en la evolución de la condición de tetrápodo. Sin embargo, no se comprende bien el motivo del desplazamiento evolutivo de la fosa nasal posterior desde la nariz hasta el labio.

Los parientes de Kenichthys pronto se establecieron en los canales y estuarios salobres y se convirtieron en los peces óseos más numerosos en todo el Devónico y la mayor parte del Carbonífero . La anatomía básica del grupo es bien conocida gracias al trabajo muy detallado sobre Eusthenopteron por Erik Jarvik en la segunda mitad del siglo XX. [33] Los huesos del techo del cráneo eran ampliamente similares a los de los primeros tetrápodos y los dientes tenían un repliegue del esmalte similar al de los laberintodontes . Las aletas pareadas tenían una estructura con huesos claramente homólogos al húmero , cúbito y radio en las aletas delanteras y al fémur , tibia y peroné en las aletas pélvicas. [34]
Existían varias familias: Rhizodontida , Canowindridae , Elpistostegidae , Megalichthyidae , Osteolepidae y Tristichopteridae . [35] La mayoría eran peces de aguas abiertas, y algunos crecían hasta alcanzar tamaños muy grandes; los especímenes adultos miden varios metros de largo. [36] Se estima que el rizodóntido Rhizodus creció hasta 7 metros (23 pies), lo que lo convierte en el pez de agua dulce más grande conocido. [37]
Aunque la mayoría de estos peces eran de aguas abiertas, un grupo, los elpistostegalianos , se adaptaron a la vida en aguas poco profundas. Desarrollaron cuerpos planos para moverse en aguas muy poco profundas, y las aletas pectorales y pélvicas asumieron el papel de los principales órganos de propulsión. La mayoría de las aletas medias desaparecieron, dejando solo una aleta caudal protocerca . Dado que las aguas poco profundas estaban sujetas a una deficiencia ocasional de oxígeno, la capacidad de respirar aire atmosférico con la vejiga natatoria se volvió cada vez más importante. [6] El espiráculo se volvió grande y prominente, lo que permitió a estos peces aspirar aire.
Los tetrápodos tienen su origen en los peces tetrapodomorfos del Devónico temprano . [38] Los tetrápodos primitivos se desarrollaron a partir de un pez tetrapodomorfo osteolépido de aletas lobuladas (sarcopterigio-crossopterigio), con un cerebro bilobulado en un cráneo aplanado . El grupo de los celacantos representa a los sarcopterigios marinos que nunca adquirieron estas adaptaciones para aguas poco profundas. Los sarcopterigios aparentemente tomaron dos líneas de descendencia diferentes y, en consecuencia, se separan en dos grupos principales: los Actinistia (incluidos los celacantos) y los Rhipidistia (que incluyen líneas extintas de peces de aletas lobuladas que evolucionaron hasta convertirse en los peces pulmonados y los tetrapodomorfos).

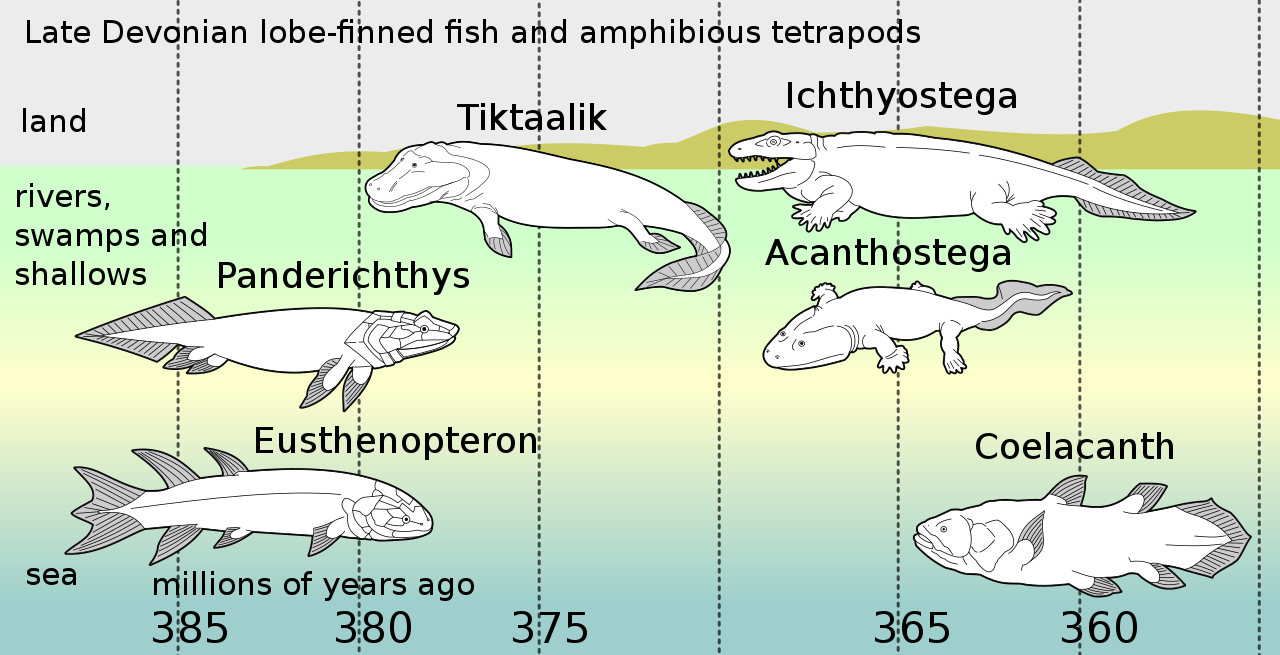
El tetrapodomorfo más antiguo conocido es Kenichthys de China, que data de alrededor de 395 millones de años. Dos de los primeros tetrapodomorfos, que datan de 380 Ma, fueron Gogonasus y Panderichthys . [39] Tenían coanas y usaban sus aletas para moverse a través de canales de marea y aguas poco profundas ahogadas con ramas muertas y plantas podridas. [40] Sus aletas podrían haber sido utilizadas para sujetarse a plantas o similares mientras acechaban a sus presas. Las características universales de los tetrápodos de las extremidades delanteras que se doblan hacia adelante desde el codo y las extremidades traseras que se doblan hacia atrás desde la rodilla pueden rastrearse plausiblemente hasta los primeros tetrápodos que vivían en aguas poco profundas. Los fósiles de huesos pélvicos de Tiktaalik muestran, si bien son representativos de los primeros tetrápodos en general, que los apéndices traseros y la locomoción propulsada por la pelvis se originaron en el agua antes de las adaptaciones terrestres. [41]
Otro indicio de que los pies y otros rasgos de los tetrápodos evolucionaron mientras los animales aún eran acuáticos es la forma en que se alimentaban. No tenían las modificaciones del cráneo y la mandíbula que les permitían tragar presas en tierra. Podían atraparlas en aguas poco profundas, al borde del agua o en tierra, pero tenían que comerlas en el agua, donde las fuerzas hidrodinámicas de la expansión de su cavidad bucal obligaban a la comida a pasar hacia el esófago. [42]
Se ha sugerido que la evolución de las extremidades de los tetrápodos a partir de las aletas en los peces de aletas lobuladas está relacionada con la expresión del gen HOXD13 o la pérdida de las proteínas actinodina 1 y actinodina 2, que están involucradas en el desarrollo de las aletas de los peces. [43] [44] Las simulaciones de robots sugieren que los circuitos nerviosos necesarios para caminar evolucionaron a partir de los nervios que gobiernan la natación, utilizando la oscilación lateral del cuerpo con las extremidades funcionando principalmente como puntos de anclaje y proporcionando un empuje limitado. [45] Este tipo de movimiento, así como los cambios en la cintura escapular son similares a los observados en el registro fósil, se pueden inducir en los bichires levantándolos fuera del agua. [46]
Un estudio de 2012 que utilizó reconstrucciones 3D de Ichthyostega concluyó que era incapaz de realizar marchas cuadrúpedas típicas . Las extremidades no podían moverse de forma alternada ya que carecían del rango de movimiento rotatorio necesario. Además, las extremidades traseras carecían de la musculatura pélvica necesaria para el movimiento terrestre impulsado por las extremidades traseras. Su método más probable de locomoción terrestre es el de "movimientos de muletas" sincrónicos, similar a los modernos saltarines del fango . [47] (Ver varios videos de saltarines del fango "caminando" muestra que se mueven empujándose hacia adelante con ambas aletas pectorales al mismo tiempo (las aletas pectorales izquierda y derecha se mueven simultáneamente, no alternativamente). Las aletas se llevan hacia adelante y se plantan; luego los hombros giran hacia atrás, avanzando el cuerpo y arrastrando la cola como un tercer punto de contacto. No hay "extremidades"/aletas traseras, y no hay una flexión significativa de la columna involucrada).
Los primeros tetrápodos probablemente evolucionaron en ambientes costeros y marinos salobres , y en hábitats de agua dulce poco profundos y pantanosos . [48] Anteriormente, los investigadores pensaban que el momento fue hacia el final del Devónico. En 2010, esta creencia fue desafiada por el descubrimiento de las huellas de tetrápodos más antiguas conocidas llamadas Zachelmie trackways , preservadas en sedimentos marinos de la costa sur de Laurasia , ahora montañas Świętokrzyskie (Santa Cruz) de Polonia. Fueron hechas durante la era Eifelian , principios del Devónico medio. Las huellas, algunas de las cuales muestran dígitos, datan de hace unos 395 millones de años, 18 millones de años antes que los fósiles de cuerpo de tetrápodo más antiguos conocidos. [49] Además, las huellas muestran que el animal era capaz de empujar sus brazos y piernas hacia adelante, un tipo de movimiento que habría sido imposible en peces tetrapodomorfos como Tiktaalik . Se estima que el animal que produjo las huellas medía hasta 2,5 metros (8,2 pies) de largo con almohadillas plantares de hasta 26 centímetros (10 pulgadas) de ancho, aunque la mayoría de las huellas tienen solo 15 centímetros (5,9 pulgadas) de ancho. [50]
Los nuevos hallazgos sugieren que los primeros tetrápodos pueden haber vivido como oportunistas en las marismas, alimentándose de animales marinos que eran arrastrados o varados por la marea. [49] Sin embargo, actualmente, los peces varados en cantidades significativas solo en ciertas épocas del año, como en la temporada de desove de la sábala ; tales varamientos no podían proporcionar un suministro significativo de alimento para los depredadores. No hay razón para suponer que los peces del Devónico fueran menos prudentes que los de hoy. [51] Según Melina Hale de la Universidad de Chicago, no todas las huellas antiguas fueron necesariamente hechas por tetrápodos primitivos, sino que también podrían haber sido creadas por parientes de los tetrápodos que usaban sus apéndices carnosos en una locomoción similar basada en el sustrato. [52] [53]
La investigación de Jennifer A. Clack y sus colegas demostró que los primeros tetrápodos, animales similares a Acanthostega , eran completamente acuáticos y no estaban adaptados a la vida en la tierra. Esto contrasta con la opinión anterior de que los peces habían invadido primero la tierra, ya sea en busca de presas (como los modernos saltarines del fango ) o para encontrar agua cuando el estanque en el que vivían se secó, y luego desarrollaron patas, pulmones, etc.
A finales del Devónico, las plantas terrestres habían estabilizado los hábitats de agua dulce, lo que permitió el desarrollo de los primeros ecosistemas de humedales , con redes alimentarias cada vez más complejas que brindaban nuevas oportunidades. Los hábitats de agua dulce no eran los únicos lugares donde encontrar agua llena de materia orgánica y vegetación densa cerca del borde del agua. También existían hábitats pantanosos como humedales poco profundos, lagunas costeras y grandes deltas de ríos salobres, y hay muchos indicios de que este es el tipo de entorno en el que evolucionaron los tetrápodos. Se han encontrado fósiles de tetrápodos primitivos en sedimentos marinos y, dado que los fósiles de tetrápodos primitivos en general se encuentran dispersos por todo el mundo, deben haberse extendido siguiendo las líneas costeras; no podrían haber vivido solo en agua dulce.
Un análisis de la Universidad de Oregon sugiere que no hay evidencia de la teoría de la "poza de agua que se encoge" (los fósiles de transición no están asociados con evidencia de charcas o estanques que se encogen) e indica que dichos animales probablemente no habrían sobrevivido a caminatas cortas entre pozos de agua agotados. [54] La nueva teoría sugiere, en cambio, que los protopulmones y protoextremidades fueron adaptaciones útiles para negociar el medio ambiente en llanuras aluviales húmedas y boscosas. [55]
Los tetrápodos del Devónico atravesaron dos grandes cuellos de botella durante lo que se conoce como la extinción del Devónico tardío ; uno al final de la etapa Frasniana y otro dos veces más grande al final de la siguiente etapa Fameniana . Estos eventos de extinciones llevaron a la desaparición de tetrápodos primitivos con características similares a los peces como Ichthyostega y sus parientes primarios más acuáticos. [56] Cuando los tetrápodos reaparecen en el registro fósil después de las extinciones del Devónico, las formas adultas están completamente adaptadas a una existencia terrestre, y las especies posteriores se adaptaron secundariamente a un estilo de vida acuático. [57]
Ahora está claro que el ancestro común de los peces óseos (Osteichthyes) tenía un pulmón primitivo que respiraba aire , que luego evolucionó hasta convertirse en una vejiga natatoria en la mayoría de los actinopterigios (peces con aletas radiadas). Esto sugiere que los crossopterigios evolucionaron en aguas cálidas y poco profundas, y que utilizaban su pulmón simple cuando el nivel de oxígeno en el agua era demasiado bajo.
Las aletas lobuladas carnosas sostenidas por huesos en lugar de aletas rígidas por radios parecen haber sido un rasgo ancestral de todos los peces óseos ( Osteichthyes ). Los ancestros con aletas lobuladas de los tetrápodos las desarrollaron aún más, mientras que los ancestros de los peces con aletas radiadas ( Actinopterygii ) desarrollaron sus aletas en una dirección diferente. El grupo más primitivo de actinopterigios, los bichiros , todavía tienen aletas frontales carnosas.
Se han descrito nueve géneros de tetrápodos del Devónico, varios de los cuales se conocen principalmente o en su totalidad a partir de material de la mandíbula inferior . Todos menos uno eran del supercontinente Laurasiático , que comprendía Europa , América del Norte y Groenlandia . La única excepción es un solo género de Gondwana , Metaxygnathus , que se ha encontrado en Australia .
El primer tetrápodo devónico identificado en Asia fue reconocido a partir de una mandíbula fósil descubierta en 2002. El tetrápodo chino Sinostega pani fue descubierto entre plantas tropicales fosilizadas y peces de aletas lobuladas en los sedimentos de arenisca roja de la región autónoma de Ningxia Hui en el noroeste de China. Este hallazgo amplió sustancialmente el área de distribución geográfica de estos animales y ha planteado nuevas preguntas sobre la distribución mundial y la gran diversidad taxonómica que alcanzaron en un tiempo relativamente corto.










Estos primeros tetrápodos no eran terrestres. Las primeras formas terrestres confirmadas se conocen a partir de los primeros depósitos del Carbonífero , unos 20 millones de años después. Aun así, es posible que hayan pasado períodos muy breves fuera del agua y que hayan usado sus patas para abrirse paso a través del barro .
Todavía se debate por qué se trasladaron a tierra en primer lugar. Una razón podría ser que los pequeños juveniles que habían completado su metamorfosis tenían lo necesario para aprovechar lo que la tierra les ofrecía. Ya adaptados a respirar aire y moverse en aguas poco profundas cerca de la tierra como protección (al igual que los peces y anfibios modernos a menudo pasan la primera parte de su vida en la relativa seguridad de aguas poco profundas como los bosques de manglares ), dos nichos muy diferentes se superponían parcialmente entre sí, con los jóvenes juveniles en la línea difusa entre ellos. Uno de ellos estaba superpoblado y era peligroso, mientras que el otro era mucho más seguro y menos abarrotado, lo que ofrecía menos competencia por los recursos. El nicho terrestre también era un lugar mucho más desafiante para los animales principalmente acuáticos, pero debido a la forma en que funcionan la evolución y la presión selectiva, los juveniles que podían aprovecharlo serían recompensados. Una vez que lograron un pequeño punto de apoyo en la tierra, gracias a sus preadaptaciones, las variaciones favorables en sus descendientes darían lugar gradualmente a una evolución y diversificación continuas.
En esa época, la abundancia de invertebrados que se arrastraban por la tierra y cerca del agua, en el suelo húmedo y en la hojarasca mojada, ofrecían una fuente de alimento. Algunos eran incluso lo suficientemente grandes como para comer pequeños tetrápodos, pero la tierra estaba libre de los peligros habituales en el agua.
Los tetrápodos, que inicialmente solo hacían incursiones tentativas en la tierra, con el tiempo se adaptaron a los entornos terrestres y pasaron períodos más largos fuera del agua. También es posible que los adultos comenzaran a pasar algún tiempo en la tierra (como sugieren las modificaciones esqueléticas en los primeros tetrápodos, como Ichthyostega ) para tomar el sol cerca de la orilla del agua [ cita requerida ] , mientras que por lo demás eran principalmente acuáticos.
Sin embargo, recientes análisis microanatómicos e histológicos de especímenes fósiles de tetrápodos encontraron que los primeros tetrápodos como Acanthostega eran completamente acuáticos, lo que sugiere que la adaptación a la tierra ocurrió más tarde. [58]
Las investigaciones de Per Ahlberg y sus colegas sugieren que las mareas podrían haber sido una fuerza impulsora de la evolución de los tetrápodos. La hipótesis propone que, a medida que "la marea retrocedía, los peces quedaron varados en entornos de pozas de marea de aguas poco profundas, donde estarían sujetos a temperaturas más altas y condiciones hipóxicas" y luego se seleccionarían peces que desarrollaran "órganos eficientes para respirar aire, así como apéndices adaptados para la navegación terrestre". [59]
Hasta la década de 1990, había una brecha de 30 millones de años en el registro fósil entre los tetrápodos del Devónico tardío y la reaparición de fósiles de tetrápodos en linajes reconocibles de anfibios del Carbonífero medio . Se la denominó " brecha de Romer ", que ahora cubre el período de hace unos 360 a 345 millones de años (la transición del Devónico al Carbonífero y principios del Misisipiense), en honor al paleontólogo que la reconoció.
Durante el "vacío" se desarrollaron las columnas vertebrales de los tetrápodos, así como las extremidades con dedos y otras adaptaciones para la vida terrestre. Las orejas , los cráneos y las columnas vertebrales también sufrieron cambios. El número de dedos en las manos y los pies se estandarizó en cinco, a medida que se extinguían los linajes con más dedos. Por lo tanto, los pocos fósiles de tetrápodos encontrados en este "vacío" son aún más apreciados por los paleontólogos porque documentan estos cambios significativos y aclaran su historia.
La transición de un pez acuático de aletas lobuladas a un anfibio que respira aire fue significativa y fundamental en la historia evolutiva de los vertebrados . Para que un organismo viviera en un entorno acuoso neutro en gravedad y luego colonizara uno que requiriera que un organismo soportara todo su peso y poseyera un mecanismo para mitigar la deshidratación, se requirieron adaptaciones o exaptaciones significativas dentro del plan corporal general , tanto en forma como en función. Eryops , un ejemplo de un animal que realizó tales adaptaciones, refinó muchos de los rasgos encontrados en sus ancestros peces. Las extremidades robustas sostenían y transportaban su cuerpo mientras estaba fuera del agua. Una columna vertebral más gruesa y fuerte impedía que su cuerpo se hundiera bajo su propio peso. Además, a través de la remodelación de los huesos vestigiales de la mandíbula de los peces, comenzó a desarrollarse un oído medio rudimentario para conectarse con el oído interno de los peces , lo que le permitió a Eryops amplificar, y así percibir mejor, el sonido transmitido por el aire .
En la etapa Viseana (mediados del Carbonífero temprano), los primeros tetrápodos se habían irradiado en al menos tres o cuatro ramas principales. Algunas de estas diferentes ramas representan los ancestros de todos los tetrápodos actuales. Esto significa que el ancestro común de todos los tetrápodos actuales probablemente vivió en el Carbonífero temprano. Bajo una definición cladística estrecha de Tetrapoda (también conocido como Crown-Tetrapoda), que solo incluye a los descendientes de este ancestro común, los tetrápodos aparecieron por primera vez en el Carbonífero. Los primeros tetrápodos reconocibles (en el sentido amplio) son representativos de los temnospóndilos (por ejemplo, Eryops ), los lepospóndilos (por ejemplo, Diplocaulus ), los antracosaurios , que eran parientes y ancestros de los amniotas , y posiblemente los bafétidos , que se cree que están relacionados con los temnospóndilos y cuyo estado como rama principal aún no se ha resuelto. Dependiendo de las autoridades que se sigan, los anfibios modernos (ranas, salamandras y cecilias ) probablemente derivan de temnospóndilos o lepospóndilos (o posiblemente de ambos, aunque ahora esta es una posición minoritaria).
Los primeros amniotas (clado de vertebrados que hoy incluye reptiles , mamíferos y aves ) son conocidos desde la primera parte del Carbonífero Tardío . En el Triásico , este grupo ya se había irradiado hacia los primeros mamíferos , tortugas y cocodrilos ( los lagartos y las aves aparecieron en el Jurásico , y las serpientes en el Cretácico ). Esto contrasta marcadamente con el grupo (posiblemente cuarto) del Carbonífero , los bafétidos , que no han dejado linajes supervivientes existentes.
Los anfibios y reptiles se vieron fuertemente afectados por el colapso de la selva tropical del Carbonífero (CRC), un evento de extinción que ocurrió hace ~307 millones de años. El período Carbonífero ha estado asociado durante mucho tiempo con pantanos espesos y húmedos y selvas tropicales húmedas. [60] Dado que las plantas forman la base de casi todos los ecosistemas de la Tierra, cualquier cambio en la distribución de las plantas siempre ha afectado a la vida animal en algún grado. El colapso repentino del vital ecosistema de la selva tropical afectó profundamente la diversidad y abundancia de los principales grupos de tetrápodos que dependían de él. [61] El CRC, que fue parte de una de las dos extinciones de plantas más devastadoras en la historia de la Tierra, fue un cambio de medio ambiente que se reforzó a sí mismo y muy rápido en el que el clima mundial se volvió mucho más seco y más frío en general (aunque se están realizando muchos trabajos nuevos para comprender mejor los cambios climáticos históricos de grano fino en la transición Carbonífero-Pérmico y cómo surgieron [62] ).
La consiguiente reducción de la población mundial de plantas, como resultado de las dificultades que encontraron las plantas para adaptarse al nuevo clima, provocó una fragmentación progresiva y el colapso de los ecosistemas de la selva tropical. Esto reforzó y aceleró aún más el colapso al reducir drásticamente la cantidad de vida animal que podían sustentar los ecosistemas en contracción en ese momento. El resultado de esta reducción animal fue un colapso en los niveles globales de dióxido de carbono, que afectó aún más a las plantas. [63] La aridez y la caída de la temperatura que resultaron de esta reducción descontrolada de la población de plantas y la disminución de un gas de efecto invernadero primario hicieron que la Tierra entrara rápidamente en una serie de intensas Edades de Hielo. [60]
Esto afectó a los anfibios en particular de varias maneras. La enorme caída del nivel del mar debido a que mayores cantidades de agua del mundo quedaron atrapadas en los glaciares afectó profundamente la distribución y el tamaño de los ecosistemas semiacuáticos que favorecían los anfibios, y el enfriamiento significativo del clima redujo aún más la cantidad de nuevo territorio favorable para los anfibios. Dado que entre las características distintivas de los anfibios se encuentran el retorno obligatorio a un cuerpo de agua para poner huevos, una piel delicada propensa a la desecación (lo que a menudo requiere que el anfibio esté relativamente cerca del agua durante toda su vida) y una reputación de ser una especie de referencia para los ecosistemas perturbados debido a la baja resiliencia resultante al cambio ecológico, [64] los anfibios fueron particularmente devastados, y los laberintodontes fueron uno de los grupos que tuvieron la peor suerte. En contraste, los reptiles, cuyos huevos amnióticos tienen una membrana que permite el intercambio de gases fuera del agua y que, por lo tanto, pueden ser puestos en la tierra, se adaptaron mejor a las nuevas condiciones. Los reptiles invadieron nuevos nichos a un ritmo más rápido y comenzaron a diversificar sus dietas, volviéndose herbívoros y carnívoros, en lugar de alimentarse exclusivamente de insectos y peces. [65] Mientras tanto, los anfibios severamente afectados simplemente no pudieron competir con los reptiles en el dominio de los nuevos nichos ecológicos, [66] y por eso se vieron obligados a pasar la antorcha evolutiva de los tetrápodos a los reptiles cada vez más exitosos y de rápida expansión.
En el período Pérmico : los primeros clados de "anfibios" (laberintodontes) incluían a los temnospóndilos y a los antracosaurios ; mientras que los clados de amniotas incluían a los saurópsidos y a los sinápsidos . Los saurópsidos finalmente evolucionarían hasta convertirse en los reptiles y aves actuales ; mientras que los sinápsidos evolucionarían hasta convertirse en los mamíferos actuales . Sin embargo, durante el Pérmico , la distinción era menos clara: la fauna de amniotas se describía típicamente como reptiles o como reptiles parecidos a mamíferos . Estos últimos (sinapsidos) fueron los animales más importantes y exitosos del Pérmico.
El final del Pérmico fue testigo de un importante cambio en la fauna durante la extinción masiva del Pérmico-Triásico , probablemente la más grave del fanerozoico . Se produjo una pérdida prolongada de especies debido a múltiples pulsos de extinción. [67] Muchos de los grupos que alguna vez fueron grandes y diversos se extinguieron o se redujeron considerablemente.
La vida en la Tierra pareció recuperarse rápidamente después de las extinciones del Pérmico, aunque esto fue principalmente en forma de taxones de desastre como el resistente Lystrosaurus . Los animales especializados que formaron ecosistemas complejos con alta biodiversidad, redes alimentarias complejas y una variedad de nichos, tardaron mucho más en recuperarse. [67] La investigación actual indica que esta larga recuperación se debió a sucesivas olas de extinción, que inhibieron la recuperación, y al estrés ambiental prolongado de los organismos que continuó hasta el Triásico Temprano. Investigaciones recientes indican que la recuperación no comenzó hasta principios del Triásico medio, 4 millones a 6 millones de años después de la extinción; [68] y algunos autores estiman que la recuperación no fue completa hasta 30 millones de años después de la extinción P-Tr, es decir, a fines del Triásico. [67]
Un pequeño grupo de reptiles, los diápsidos , comenzó a diversificarse durante el Triásico, en particular los dinosaurios . A finales del Mesozoico, los grandes grupos de laberintodontes que aparecieron por primera vez durante el Paleozoico, como los temnospóndilos y los anfibios similares a reptiles , se habían extinguido. Todos los grupos principales actuales de saurópsidos evolucionaron durante el Mesozoico, y las aves aparecieron por primera vez en el Jurásico como un clado derivado de los dinosaurios terópodos . Muchos grupos de sinápsidos como los anomodontes y los terocéfalos que alguna vez comprendieron la fauna terrestre dominante del Pérmico también se extinguieron durante el Mesozoico; sin embargo, durante el Triásico, un grupo ( Cynodontia ) dio lugar al taxón descendiente Mammalia , que sobrevivió durante el Mesozoico para luego diversificarse durante el Cenozoico.
La era Cenozoica comenzó con el fin de la era Mesozoica y la época Cretácica , y continúa hasta nuestros días. El comienzo de la era Cenozoica estuvo marcado por el evento de extinción del Cretácico-Paleógeno durante el cual se extinguieron todos los dinosaurios no aviares . La era Cenozoica a veces se denomina la "Era de los Mamíferos ".
Durante el Mesozoico, el mamífero prototípico era un pequeño insectívoro nocturno , algo así como una musaraña arbórea . Debido a sus hábitos nocturnos , la mayoría de los mamíferos perdieron su visión del color y mejoraron enormemente su sentido del olfato y el oído . Todos los mamíferos de la actualidad están moldeados por este origen. Los primates y algunos marsupiales australianos volvieron a desarrollar posteriormente la visión del color.
Durante el Paleoceno y el Eoceno , la mayoría de los mamíferos eran pequeños (menos de 20 kg). El enfriamiento del clima en el Oligoceno y el Mioceno y la expansión de los pastizales favorecieron la evolución de especies de mamíferos más grandes.
Las aves corredoras corren y los pingüinos nadan y se balancean, pero la mayoría de las aves son bastante pequeñas y pueden volar. Algunas aves utilizan su capacidad de volar para completar migraciones épicas que cruzan el mundo , mientras que otras, como las fragatas, sobrevuelan los océanos durante meses.
Los murciélagos también han emprendido el vuelo y, junto con los cetáceos, han desarrollado la ecolocalización o sonar .
Las ballenas , las focas , los manatíes y las nutrias marinas han regresado al océano y a un estilo de vida acuático .
Grandes manadas de ungulados rumiantes pueblan los pastizales y los bosques . Los carnívoros han evolucionado para mantener a raya las poblaciones de animales en manada.
Tras la gran renovación faunística de finales del Mesozoico, sólo quedaron siete grupos de tetrápodos, uno de los cuales, los Choristodera , se extinguió hace 11 millones de años por razones desconocidas. Los otros seis que persisten en la actualidad también incluyen muchos miembros extintos:
Un pariente aún más cercano era
Panderichthys
, que incluso tenía una coana. Estos peces usaban sus aletas como remos en hábitats de aguas poco profundas repletos de plantas y detritos.
{kind=link}