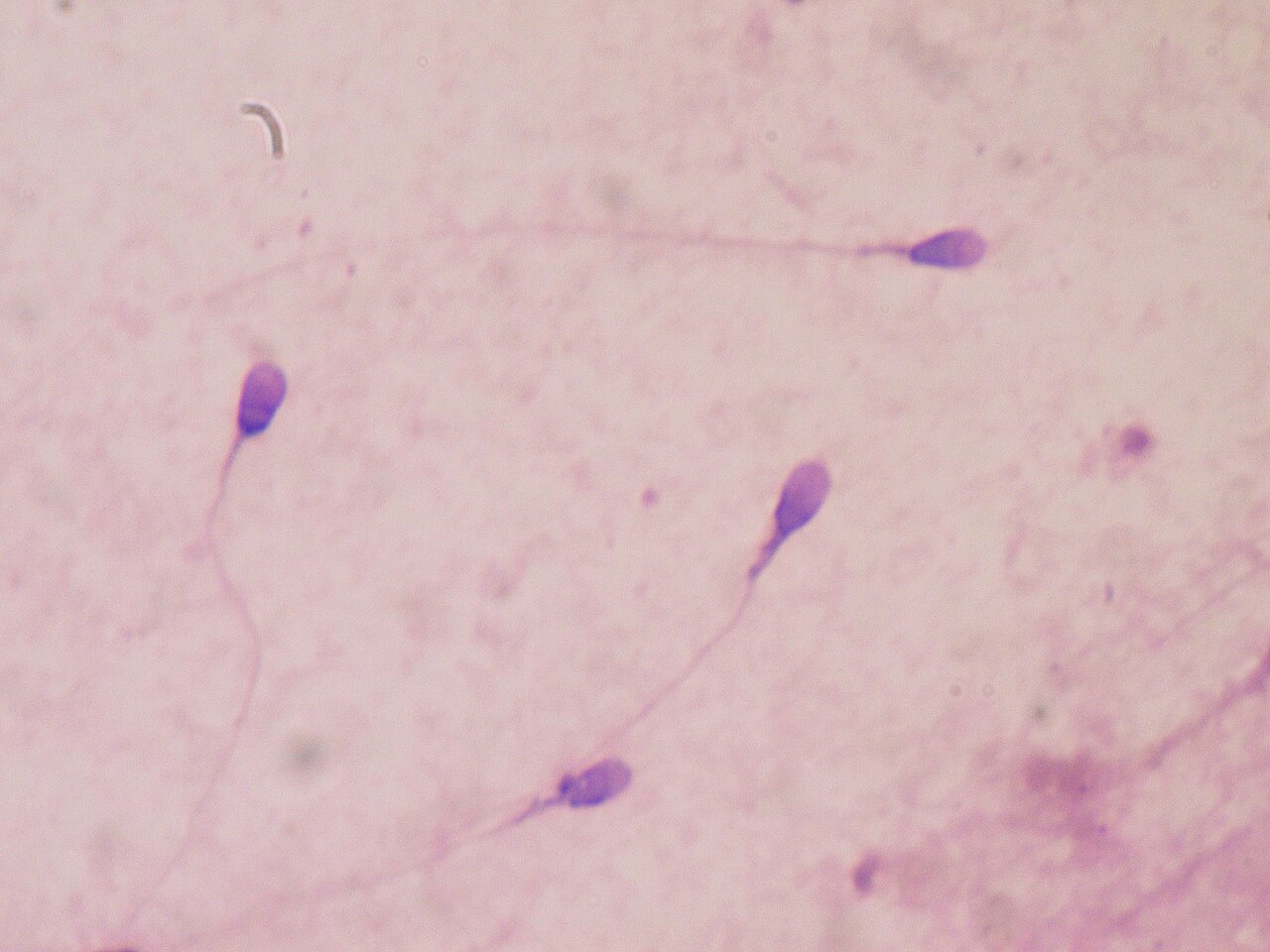

La competencia espermática es el proceso competitivo entre los espermatozoides de dos o más machos diferentes para fertilizar el mismo óvulo [1] durante la reproducción sexual . La competencia puede ocurrir cuando las hembras tienen múltiples parejas potenciales de apareamiento. Una mayor elección y variedad de parejas aumenta la posibilidad de que una hembra produzca descendencia más viable. [2] Sin embargo, múltiples parejas para una hembra significa que cada macho individual tiene menos posibilidades de producir descendencia. La competencia espermática es una presión evolutiva sobre los machos y ha llevado al desarrollo de adaptaciones para aumentar la posibilidad de éxito reproductivo de los machos . [3] La competencia espermática da como resultado un conflicto sexual entre machos y hembras. [2] Los machos han desarrollado varias tácticas defensivas que incluyen: protección de la pareja, tapones de apareamiento y liberación de sustancias seminales tóxicas para reducir las tendencias de las hembras a volver a aparearse para hacer frente a la competencia espermática. [4] Las tácticas ofensivas de la competencia espermática implican la interferencia directa de un macho en el éxito reproductivo de otro macho, por ejemplo, mediante la protección de la pareja o eliminando físicamente el esperma de otro macho antes de aparearse con una hembra. [5] [6] Para un ejemplo, véase Gryllus bimaculatus .
La competencia de espermatozoides se compara a menudo con tener boletos en una rifa ; un macho tiene más posibilidades de ganar (es decir, engendrar descendencia) cuanto más boletos tenga (es decir, cuanto más espermatozoides use para inseminar a una hembra). Sin embargo, los espermatozoides no son libres de producir, [7] y, como tal, se predice que los machos producirán espermatozoides de un tamaño y una cantidad que maximizarán su éxito en la competencia de espermatozoides. Al producir muchos espermatozoides, los machos pueden comprar más "boletos de rifa", y se cree que la selección de numerosos espermatozoides ha contribuido a la evolución de la anisogamia con espermatozoides muy pequeños (debido al equilibrio energético entre el tamaño y la cantidad de espermatozoides). [8] Alternativamente, un macho puede desarrollar espermatozoides más rápidos para permitir que sus espermatozoides alcancen y fertilicen primero el óvulo de la hembra . Se han documentado docenas de adaptaciones en los machos que los ayudan a tener éxito en la competencia de espermatozoides.
.JPG/1280px-Neolamprologus_pulcher_(Wroclaw_zoo).JPG)
La vigilancia de la pareja es un rasgo de comportamiento defensivo que se produce en respuesta a la competencia de los espermatozoides; los machos tratan de impedir que otros machos se acerquen a la hembra (y/o viceversa), impidiendo así que su pareja participe en más cópulas . [2] La vigilancia de la pareja precopulatoria y poscopulatoria se produce en insectos, lagartijas, aves y primates. La vigilancia de la pareja también existe en la especie de pez Neolamprologus pulcher , ya que algunos machos intentan "esconder" apareamientos con hembras en el territorio de otros machos. En estos casos, los machos protegen a su hembra manteniéndola lo suficientemente cerca como para que si un macho oponente aparece en su territorio pueda luchar contra el macho rival, lo que evitará que la hembra participe en cópulas extraparejas con el macho rival. [9]
Los organismos con sistemas de apareamiento poligínicos están controlados por un macho dominante. En este tipo de sistema de apareamiento, el macho puede aparearse con más de una hembra en una comunidad. [10] Los machos dominantes reinarán sobre la comunidad hasta que otro pretendiente se presente y lo derroque. [10] El macho dominante actual defenderá su título como macho dominante y también defenderá a las hembras con las que se aparee y a las crías que engendre. El elefante marino entra en esta categoría, ya que puede participar en combates violentos y sangrientos para proteger a su comunidad y defender su título de macho alfa. [11] Si el macho alfa es derrocado de alguna manera por el recién llegado, lo más probable es que sus hijos mueran y el nuevo macho alfa comience de nuevo con las hembras del grupo para que su linaje pueda transmitirse. [12]
La protección estratégica de la pareja se produce cuando el macho sólo protege a la hembra durante sus períodos fértiles. Esta estrategia puede ser más eficaz porque puede permitir al macho ejercer tanto la paternidad extrapareja como la paternidad dentro de la pareja. [13] Esto también se debe a que es energéticamente eficiente para el macho proteger a su pareja en este momento. Se gasta mucha energía cuando un macho protege a su pareja. Por ejemplo, en los sistemas de protección de pareja polígamos, el coste energético de los machos es defender su título de macho alfa de su comunidad. [11] Las peleas son muy costosas en cuanto a la cantidad de energía utilizada para proteger a su pareja. Estos combates pueden ocurrir más de una vez, lo que afecta al bienestar físico del macho. Otro coste de la protección de la pareja en este tipo de sistema de apareamiento es el posible aumento de la propagación de enfermedades. [14] Si un macho tiene una ETS, puede transmitirla a las hembras con las que está copulando, lo que puede dar lugar a un agotamiento del harén. Esto supondría un coste energético para ambos sexos, ya que en lugar de utilizar la energía para la reproducción, la están redirigiendo hacia la eliminación de esta enfermedad. Algunas hembras también se benefician de la poligamia porque las cópulas de parejas adicionales en las hembras aumentan la diversidad genética con la comunidad de esa especie. [15] Esto ocurre porque el macho no es capaz de vigilar a todas las hembras y algunas se volverán promiscuas. Con el tiempo, el macho no tendrá una nutrición adecuada, lo que lo hará incapaz de producir esperma. [16] Por ejemplo, los anfípodos macho agotarán sus reservas de glucógeno y triglicéridos solo para reponerlas después de que el macho haya terminado de proteger a esa pareja. [17] Además, si la cantidad de ingesta de energía no es igual a la energía gastada, esto podría ser potencialmente fatal para el macho. Los machos pueden incluso tener que viajar largas distancias durante la temporada de reproducción para encontrar una hembra, lo que drena significativamente su suministro de energía. Se realizaron estudios para comparar el costo de la búsqueda de alimento de los peces que migran y los animales que son residenciales. Los estudios concluyeron que los peces que eran residentes tenían estómagos más llenos que contenían presas de mayor calidad en comparación con sus contrapartes migrantes. [18] Con todos estos costos de energía que conlleva la protección de una pareja, el momento es crucial para que el macho pueda utilizar la cantidad mínima de energía. Es por eso que es más eficiente para los machos elegir una pareja durante sus períodos fértiles. [13] Además, los machos tendrán más probabilidades de proteger a su pareja cuando hay una alta densidad de machos en la proximidad. [2]A veces, los organismos dedican todo este tiempo y planificación a cortejar a una pareja para copular y ella puede ni siquiera estar interesada. Existe el riesgo de que se cometa algún tipo de engaño, ya que un macho rival puede cortejar con éxito a la hembra, algo que el macho que la cortejó originalmente no pudo hacer. [19]
Sin embargo, existen beneficios asociados con la protección de la pareja. En un sistema de protección de la pareja, ambas partes, macho y hembra, pueden beneficiarse directa e indirectamente de esto. Por ejemplo, las hembras pueden beneficiarse indirectamente de ser protegidas por un compañero. [20] Las hembras pueden apreciar una disminución en la depredación y el acoso de otros machos mientras pueden observar a su contraparte masculina. [20] Esto le permitirá reconocer rasgos particulares que le parezcan ideales para poder encontrar otro macho que emule esas cualidades. En las relaciones poligínicas, el macho dominante de la comunidad se beneficia porque tiene el mejor éxito de fertilización. [12] Las comunidades pueden incluir de 30 a 100 hembras y, en comparación con los otros machos, aumentarán en gran medida sus posibilidades de éxito de apareamiento. [11]
Los machos que han cortejado con éxito a una pareja potencial intentarán mantenerla fuera de la vista de otros machos antes de la cópula. Una forma en que los organismos logran esto es moviendo a la hembra a una nueva ubicación. Algunas mariposas, después de seducir a la hembra, la recogen y la alejan volando de la proximidad de los machos potenciales. [21] En otros insectos, los machos liberan una feromona para hacer que su pareja sea poco atractiva para otros machos, o para enmascarar su olor por completo. [21] El macho de ciertas especies de grillos cortejará a una hembra en voz alta, hasta que ella acepte su gesto, cuando de repente se queda en silencio. [22] Algunos insectos, antes de aparearse, asumirán posiciones de tándem con su pareja o se posicionarán de una manera que evite que otros machos intenten aparearse con esa hembra. [21] La mariposa damero macho ha desarrollado un método inteligente para atraer y proteger a una pareja. Se situará cerca de un área que posea recursos valiosos que la hembra necesita. Luego ahuyentará a cualquier macho que se acerque y esto aumentará enormemente sus posibilidades de copular con cualquier hembra que llegue a esa zona. [23]
En la vigilancia de la pareja después de la cópula, los machos intentan evitar que otros machos se apareen con la hembra con la que ya se han apareado. Por ejemplo, los milpiés machos de Costa Rica se montan sobre la espalda de su pareja para hacerles saber a los demás machos que está preñada. [24] Los escarabajos japoneses adoptan una posición de tándem con la hembra después de la cópula. [25] Esto puede durar varias horas, lo que le permite alejar a los machos rivales, lo que da a su esperma una gran posibilidad de fertilizar el óvulo de esa hembra. Estos y otros tipos de métodos hacen que el macho defienda a su pareja. Se sabe que los elefantes marinos participan en sangrientas batallas para conservar su título de macho dominante y poder aparearse con todas las hembras de su comunidad. [11]

.jpg/1280px-Drosophila_melanogaster_-_side_(aka).jpg)
Los tapones copulatorios se observan con frecuencia en insectos, reptiles, algunos mamíferos y arañas. [2] Los tapones copulatorios se insertan inmediatamente después de que un macho copula con una hembra, lo que reduce la posibilidad de fertilización por cópulas posteriores de otro macho, al bloquear físicamente la transferencia de esperma. [2] Los tapones de apareamiento de abejorros , además de proporcionar una barrera física para futuras cópulas, contienen ácido linoleico , que reduce las tendencias de re-apareamiento de las hembras. [26] Una especie de Drosophila del desierto de Sonora , Drosophila mettleri , utiliza tapones copulatorios para permitir que los machos controlen el espacio de reserva de esperma que las hembras tienen disponible. Este comportamiento asegura a los machos un mayor éxito de apareamiento a expensas del control de los espermatozoides por parte de las hembras (selección de espermatozoides).
De manera similar, los machos de Drosophila melanogaster liberan fluidos seminales tóxicos, conocidos como ACP (proteínas de glándulas accesorias), desde sus glándulas accesorias para impedir que la hembra participe en futuras cópulas. [27] Estas sustancias actúan como un antiafrodisíaco provocando un rechazo de las cópulas posteriores, y también estimulan la ovulación y la ovogénesis. [5] Las proteínas seminales pueden tener una fuerte influencia en la reproducción, suficiente para manipular el comportamiento y la fisiología de las hembras. [28]
Otra estrategia, conocida como partición de esperma, ocurre cuando los machos conservan su suministro limitado de esperma reduciendo la cantidad de esperma expulsado. [2] En Drosophila , la cantidad de eyaculación durante las cópulas secuenciales se reduce; esto da como resultado reservas de esperma femenino medio llenas después de un solo evento copulatorio, pero permite al macho aparearse con un mayor número de hembras sin agotar su suministro de esperma. [2] Para facilitar la partición de esperma, algunos machos han desarrollado formas complejas de almacenar y entregar su esperma. [29] En el lábrido de cabeza azul, Thalassoma bifasciatum , el conducto de esperma está seccionado en varias cámaras pequeñas que están rodeadas por un músculo que permite al macho regular la cantidad de esperma que se libera en un evento copulatorio. [30]
Una estrategia común entre los insectos es la de los machos que participan en cópulas prolongadas. Al participar en cópulas prolongadas, el macho tiene una mayor oportunidad de colocar más esperma dentro del tracto reproductivo de la hembra y evitar que esta copule con otros machos. [31]
Se ha descubierto que algunos machos de mollies ( Poecilia ) han desarrollado señales sociales engañosas para combatir la competencia de espermatozoides. Los machos focales dirigirán su atención sexual hacia hembras que normalmente no son las preferidas cuando haya una audiencia de otros machos. Esto alienta a los machos que están mirando a intentar aparearse con la hembra no preferida. Esto se hace en un intento de disminuir los intentos de apareamiento con la hembra que el macho focal prefiere, disminuyendo así la competencia de espermatozoides. [32]
La conducta de adaptación ofensiva se diferencia de la conducta defensiva porque implica un intento de arruinar las posibilidades de que otro macho tenga éxito en la cópula al participar en un acto que intenta terminar con el éxito de fertilización del macho anterior. [5] Esta conducta ofensiva se ve facilitada por la presencia de ciertos rasgos, que se denominan armamentos. [33] Un ejemplo de armamento son las astas . Además, la presencia de un rasgo ofensivo a veces sirve como señal de estatus . La mera exhibición de un armamento puede ser suficiente para ahuyentar a la competencia sin entrar en una pelea, ahorrando así energía. [33] Un macho en el lado ofensivo de la protección de la pareja puede terminar con las posibilidades del macho guardián de una inseminación exitosa peleándose con el macho guardián para obtener acceso a la hembra. [2] En Drosophila , los machos liberan fluidos seminales que contienen toxinas adicionales como feromonas y enzimas modificadas que son secretadas por sus glándulas accesorias destinadas a destruir el esperma que ya ha llegado al tracto reproductivo de la hembra desde una cópula reciente. [5] Sin embargo, esto resultó ser incorrecto porque el fluido seminal de Drosophila melanogaster en realidad puede proteger el esperma de otros machos. [34] Basándose en la idea de la " última precedencia masculina ", algunos machos pueden eliminar el esperma de machos anteriores eyaculando nuevo esperma en la hembra; obstaculizando las oportunidades de inseminación exitosa del macho anterior. [35] Un ejemplo de este comportamiento se ve en las especies de escarabajos Carabus insulicola y Onymacris unguicularis . En el primero, el segundo macho que se aparea con una hembra puede usar sus genitales en forma de gancho para desalojar el espermatóforo depositado por el macho anterior para aumentar sus posibilidades de fertilizar la mayor cantidad de huevos. [36] En la última especie, el espermatóforo del segundo macho funciona esencialmente para expulsar el espermatóforo del último macho fuera del cuerpo de la hembra, aunque se ha demostrado que no todos los espermatozoides anteriores se eliminan por completo. [37]
La "hipótesis del buen esperma" es muy común en los sistemas de apareamiento poliándrico . [38] La "hipótesis del buen esperma" sugiere que la composición genética de un macho determinará el nivel de su competitividad en la competencia de esperma. [38] Cuando un macho tiene "buen esperma" es capaz de engendrar más crías viables que los machos que no tienen los genes del "buen esperma". [38] Las hembras pueden seleccionar machos que tienen estos genes superiores de "buen esperma" porque significa que su descendencia será más viable y heredará los genes del "buen esperma", lo que aumentará sus niveles de aptitud cuando sus espermatozoides compitan. [39]
Los estudios muestran que, además de la composición genética del macho, la competitividad de los espermatozoides en la competición espermática depende de otros factores. La ingesta dietética del macho también afectará a la competición espermática. Una dieta adecuada que consista en mayores cantidades de dieta y, a veces, una proporción más específica en ciertas especies optimizará el número y la fertilidad de los espermatozoides. Se probaron las cantidades de ingesta de proteínas y carbohidratos para determinar sus efectos sobre la producción y la calidad de los espermatozoides en moscas de la fruta adultas (Diptera: Tephritidae). Los estudios mostraron que estas moscas necesitan ingerir constantemente carbohidratos y agua para sobrevivir, pero también se requieren proteínas para alcanzar la madurez sexual. [40] Además, se ha demostrado que la dieta del macho de la mosca de la fruta mediterránea afecta al éxito del apareamiento, la duración de la cópula, la transferencia de esperma y la participación del macho en los leks. [41] Todos estos requieren una buena dieta con nutrientes para la producción adecuada de gametos, así como energía para las actividades, que incluyen la participación en los leks.
Además, se demostró que las cantidades de proteínas y carbohidratos tienen un efecto sobre la producción de esperma y la fertilidad en la cucaracha moteada. Se utilizaron dietas holídicas que permitieron tomar mediciones específicas de proteínas y carbohidratos, lo que le dio credibilidad. Se vio una correlación directa en el número de espermatozoides y la ingesta total de alimentos. Más específicamente, la producción óptima de esperma se midió en una proporción de proteínas a carbohidratos de 1:2. La fertilidad de los espermatozoides fue mejor en una proporción similar de proteínas a carbohidratos de 1:2. Esta estrecha alineación influye en gran medida en la determinación de la fertilidad masculina en Nauphoeta cinerea . [42] Sorprendentemente, la viabilidad de los espermatozoides no se vio afectada por ningún cambio en la dieta o las proporciones de la dieta. Se plantea la hipótesis de que la viabilidad de los espermatozoides se ve más afectada por la composición genética, como en la "hipótesis del buen esperma". Estas proporciones y resultados no son consistentes con muchas otras especies e incluso entran en conflicto con algunas. Parece que no puede haber ninguna conclusión sobre qué tipo de dieta se necesita para influir positivamente en la competencia de los espermatozoides, sino que más bien debe entenderse que las diferentes dietas juegan un papel en la determinación de la competencia de los espermatozoides en la elección de pareja .
Una respuesta evolutiva a la competencia espermática es la variedad en la morfología del pene de muchas especies. [43] [44] Por ejemplo, la forma del pene humano puede haber sido moldeada selectivamente por la competencia espermática. [45] El pene humano puede haber sido seleccionado para desplazar los fluidos seminales implantados en el tracto reproductivo femenino por un macho rival. [45] Específicamente, la forma de la cresta coronal puede promover el desplazamiento del fluido seminal de un apareamiento anterior [46] a través de la acción de empuje durante las relaciones sexuales . [45] Un estudio de 2003 por Gordon G. Gallup y colegas concluyó que un propósito evolutivo del movimiento de empuje característico de las relaciones sexuales intensas es que el pene "succione" el semen de otro hombre antes de depositar el suyo. [47]
La evolución para aumentar el volumen de eyaculación en presencia de competencia de espermatozoides tiene una consecuencia en el tamaño de los testículos . Los testículos grandes pueden producir más espermatozoides necesarios para eyaculados más grandes, y se pueden encontrar en todo el reino animal cuando se produce competencia de espermatozoides. [48] Se ha documentado que los machos con testículos más grandes logran mayores tasas de éxito reproductivo que los machos con testículos más pequeños en las ardillas listadas de pino amarillo macho . [48] En los peces cíclidos, se ha descubierto que una mayor competencia de espermatozoides puede conducir a una mayor cantidad de espermatozoides, tamaños de células espermáticas y velocidades de nado de los espermatozoides. [49]
En algunos insectos y arañas, por ejemplo , Nephila fenstrate , el órgano copulador masculino se rompe o se arranca al final de la cópula y permanece dentro de la hembra para servir como tapón copulador. [50] Se cree que estos genitales rotos son una respuesta evolutiva a la competencia de los espermatozoides. [50] Este daño a los genitales masculinos significa que estos machos solo pueden aparearse una vez. [51]
Los factores femeninos pueden influir en el resultado de la competencia espermática a través de un proceso conocido como "elección espermática". [52] Las proteínas presentes en el tracto reproductivo femenino o en la superficie del óvulo pueden influir en qué espermatozoide logra fertilizar el óvulo. [52] Durante la elección espermática, las hembras pueden discriminar y utilizar de forma diferente el esperma de diferentes machos. Un caso en el que se sabe que esto ocurre es la endogamia; las hembras utilizarán preferentemente el esperma de un macho con un parentesco más lejano que el de un pariente cercano. [52]
La endogamia suele tener consecuencias negativas para la aptitud ( depresión endogámica ) y, como resultado, las especies han desarrollado mecanismos para evitarla. Se considera que la depresión endogámica se debe en gran medida a la expresión de mutaciones recesivas deletéreas homocigóticas . [53] El cruzamiento entre individuos no relacionados suele conducir al enmascaramiento de mutaciones recesivas deletéreas en la progenie. [54]
Se han descrito numerosos mecanismos de evitación de la endogamia que operan antes del apareamiento. Sin embargo, los mecanismos de evitación de la endogamia que operan después de la cópula son menos conocidos. En los guppies , se produce un mecanismo de evitación de la endogamia posterior a la cópula basado en la competencia entre los espermatozoides de machos rivales para lograr la fertilización. [55] En las competencias entre los espermatozoides de un macho no emparentado y de un macho hermano completo, se observó un sesgo significativo en la paternidad hacia el macho no emparentado. [55]
Los experimentos de fertilización in vitro en ratones proporcionaron evidencia de selección de espermatozoides a nivel gamético. [56] Cuando se mezclaron espermatozoides de machos hermanos y no hermanos, se observó un sesgo de fertilización hacia los espermatozoides de los machos no hermanos. Los resultados se interpretaron como una selección de espermatozoides impulsada por los óvulos en contra de los espermatozoides relacionados.
Las moscas de la fruta hembras ( Drosophila melanogaster ) se aparearon con machos de cuatro grados diferentes de parentesco genético en experimentos de competencia. [57] La capacidad competitiva de los espermatozoides se correlacionó negativamente con el parentesco.
Las hembras de grillo ( Teleogrillus oceanicus ) parecen utilizar mecanismos poscopulatorios para evitar producir crías endogámicas. Cuando se aparean con un hermano y un macho no emparentado, las hembras predisponen la paternidad hacia el macho no emparentado. [58]

Se ha descubierto que debido a la elección femenina (ver selección sexual ), la morfología de los espermatozoides en muchas especies ocurre en muchas variaciones para acomodarse o combatir (ver conflicto sexual ) la morfología y fisiología del tracto reproductivo femenino . [59] [60] [61] Sin embargo, es difícil entender la interacción entre la forma y estructura reproductiva femenina y masculina que ocurre dentro del tracto reproductivo femenino después del apareamiento que permite la competencia de los espermatozoides. Las hembras poliándricas se aparean con muchos compañeros masculinos. [62] Las hembras de muchas especies de artrópodos , moluscos y otros filos tienen un órgano especializado de almacenamiento de esperma llamado espermateca en el que los espermatozoides de diferentes machos a veces compiten por un mayor éxito reproductivo. [60] Se sabe que las especies de grillos, específicamente Gryllus bimaculatus , exhiben selección sexual poliándrica. Los machos invertirán más en la eyaculación cuando los competidores están en el entorno inmediato de la hembra.
Existen evidencias que ilustran la capacidad de los espermatozoides genéticamente similares para cooperar de manera de asegurar la supervivencia de sus contrapartes, asegurando así la implementación de sus genotipos hacia la fertilización. La cooperación confiere una ventaja competitiva por varios medios, algunos de los cuales incluyen la incapacitación de otros espermatozoides competidores y la agregación de espermatozoides genéticamente similares en estructuras que promueven la navegación efectiva del tracto reproductivo femenino y, por lo tanto, mejoran la capacidad de fertilización. Tales características conducen a adaptaciones morfológicas que se adaptan a los propósitos de los métodos cooperativos durante la competencia. Por ejemplo, los espermatozoides que posee el ratón de campo ( Apodemus sylvaticus ) poseen un gancho apical que se utiliza para unirse a otros espermatozoides para formar trenes móviles que mejoran la motilidad a través del tracto reproductivo femenino. [63] Los espermatozoides que no se incorporan a los trenes móviles tienen menos probabilidades de participar en la fertilización. Otra evidencia sugiere que no hay vínculo entre la competencia de los espermatozoides y la morfología del gancho de los espermatozoides. [64]
La selección para producir más esperma también puede seleccionar la evolución de testículos más grandes . Las relaciones entre especies entre la frecuencia de apareamiento múltiple por parte de las hembras y el tamaño de los testículos de los machos están bien documentadas en muchos grupos de animales. Por ejemplo, entre los primates, las gorilas hembras son relativamente monógamas, por lo que las gorilas tienen testículos más pequeños que los humanos , que a su vez tienen testículos más pequeños que los altamente promiscuos bonobos . [65] Los chimpancés machos que viven en una comunidad estructurada de varios machos y varias hembras, tienen testículos grandes para producir más esperma, lo que les da mejores probabilidades de fertilizar a la hembra. Mientras que una comunidad de gorilas consta de un macho alfa y dos o tres hembras; cuando las gorilas hembras están listas para aparearse, normalmente solo el macho alfa es su pareja.
En cuanto al dimorfismo sexual entre los primates, los humanos se encuentran en un grupo intermedio con diferencias moderadas en el tamaño corporal, pero con testículos relativamente grandes. Este es un patrón típico de los primates, en el que varios machos y hembras viven juntos en un grupo y el macho se enfrenta a un número intermedio de desafíos por parte de otros machos en comparación con la poligamia y la monogamia exclusivas , pero con una competencia frecuente de espermatozoides. [66]
Otros medios de competencia de los espermatozoides podrían incluir la mejora del propio esperma o de sus materiales de empaquetamiento ( espermatóforo ). [67]
El macho de la libélula de alas negras constituye un ejemplo sorprendente de adaptación a la competencia de espermatozoides. Se sabe que las hembras de libélulas de alas negras se aparean con varios machos en el lapso de unas pocas horas y, por lo tanto, poseen un receptáculo conocido como espermateca que almacena el esperma. Durante el proceso de apareamiento, el macho de la libélula bombea su abdomen hacia arriba y hacia abajo utilizando su pene especialmente adaptado que actúa como un cepillo para eliminar el esperma de otro macho. Este método resulta bastante exitoso y se sabe que el macho de la libélula elimina el 90-100 por ciento del esperma competidor. [68]

Se ha observado una estrategia similar en el acentor común , un pequeño pájaro. Antes de aparearse con la hembra poliándrica , el acentor común macho picotea la cloaca de la hembra para extraer el esperma del pretendiente macho anterior. [69]
En la mosca Dryomyza anilis , las hembras se aparean con varios machos. Es beneficioso para el macho intentar ser el último en aparearse con una determinada hembra. [70] Esto se debe a que parece haber un aumento porcentual acumulativo en la fertilización para el último macho, de modo que los huevos puestos en la última oviposición son los más exitosos.
En 1996 surgió la idea de que en algunas especies, incluidos los humanos, una fracción significativa de los espermatozoides no puede fecundar el óvulo; más bien, se teorizó que estos espermatozoides evitaban que los espermatozoides de otros machos llegaran al óvulo, por ejemplo matándolos con enzimas o bloqueando su acceso. Este tipo de especialización de los espermatozoides se conoció popularmente como "espermatozoides kamikaze" o "espermatozoides asesinos", pero la mayoría de los estudios de seguimiento de esta noción popularizada no han podido confirmar los artículos iniciales sobre el tema. [71] [6] Si bien actualmente también hay poca evidencia de espermatozoides asesinos en cualquier animal no humano [72] ciertos caracoles tienen una forma de espermatozoide infértil ("parasperma") que contiene lisozimas , lo que lleva a la especulación de que podrían ser capaces de degradar el esperma de un rival. [73]
En la avispa parasitoide Nasonia vitripennis , las hembras apareadas pueden elegir si ponen o no un huevo fertilizado (que se convierte en una hija) o un huevo no fertilizado (que se convierte en un hijo), por lo que las hembras sufren un costo por el apareamiento, ya que los apareamientos repetidos limitan su capacidad para asignar sexo a su descendencia. El comportamiento de estos espermatozoides kamikaze se conoce en la literatura académica como "bloqueo de espermatozoides", utilizando el baloncesto como metáfora. [74]
La competencia entre los espermatozoides ha dado lugar a otras adaptaciones, como eyaculados de mayor tamaño , cópulas prolongadas , deposición de un tapón copulatorio para impedir que la hembra se vuelva a aparear o la aplicación de feromonas que reducen el atractivo de la hembra. La adaptación de los rasgos de los espermatozoides, como la longitud, la viabilidad y la velocidad, podría verse limitada por la influencia del ADN citoplasmático (por ejemplo, el ADN mitocondrial ); [75] el ADN mitocondrial se hereda únicamente de la madre y se cree que esto podría representar una limitación en la evolución de los espermatozoides.
{{cite journal}}: CS1 maint: varios nombres: lista de autores ( enlace )