Los krill (eufáusidos) [1] ( sg .: krill) son crustáceos pequeños y exclusivamente marinos del orden Euphausiacea , que se encuentran en todos los océanos del mundo. [2] El nombre "krill" proviene de la palabra noruega krill , que significa "pequeños alevines de pescado", [3] que también se atribuye a menudo a especies de peces.
El krill se considera un importante eslabón del nivel trófico cercano al fondo de la cadena alimentaria . Se alimenta de fitoplancton y, en menor medida, de zooplancton , y también es la principal fuente de alimento para muchos animales más grandes. En el océano Austral , una especie, el krill antártico , representa una biomasa estimada de alrededor de 379 millones de toneladas , [4] lo que la convierte en una de las especies con mayor biomasa total. Más de la mitad de esta biomasa es consumida por ballenas, focas , pingüinos, aves marinas, calamares y peces cada año. La mayoría de las especies de krill muestran grandes migraciones verticales diarias , proporcionando alimento a los depredadores cerca de la superficie durante la noche y en aguas más profundas durante el día.
El krill se pesca comercialmente en el océano Austral y en las aguas que rodean Japón. La cosecha mundial total asciende a 150.000–200.000 toneladas anuales, principalmente del mar de Scotia . La mayor parte de la captura de krill se utiliza para la acuicultura y la alimentación de acuarios , como cebo en la pesca deportiva o en la industria farmacéutica. El krill también se utiliza para el consumo humano en varios países. Se le conoce como okiami (オキアミ) en Japón y como camarones en España y Filipinas. En Filipinas, también se le llama alamang y se utiliza para hacer una pasta salada llamada bagoong .
El orden Euphausiacea comprende dos familias . La más abundante Euphausiidae contiene 10 géneros diferentes con un total de 85 especies. De estos, el género Euphausia es el más grande, con 31 especies. [5] La familia menos conocida, Bentheuphausiidae , tiene solo una especie , Bentheuphausia amblyops , un krill batipelágico que vive en aguas profundas por debajo de los 1000 m (3300 pies). Se considera la especie de krill existente más primitiva. [6]
A partir de 2013 [actualizar], se cree que el orden Euphausiacea es monofilético debido a varias características morfológicas únicas conservadas ( autapomorfía ), como sus branquias filamentosas desnudas y toracópodos delgados [10] y por estudios moleculares. [11] [12] [13]
Ha habido muchas teorías sobre la ubicación del orden Euphausiacea. Desde la primera descripción de Thysanopode tricuspide por Henri Milne-Edwards en 1830, la similitud de sus toracópodos birrames había llevado a los zoólogos a agrupar a los eufáusidos y a los misidáceos en el orden Schizopoda , que fue dividido por Johan Erik Vesti Boas en 1883 en dos órdenes separados. [14] Más tarde, William Thomas Calman (1904) clasificó a los misidáceos en el superorden Peracarida y a los eufáusidos en el superorden Eucarida , aunque incluso hasta la década de 1930 se defendía el orden Schizopoda. [10] Más tarde también se propuso que el orden Euphausiacea debería agruparse con los Penaeidae (familia de los camarones) en los Decapoda basándose en similitudes de desarrollo, como señalaron Robert Gurney e Isabella Gordon . [15] [16] La razón de este debate es que el krill comparte algunas características morfológicas de los decápodos y otras de los misidáceos. [10]
Los estudios moleculares no los han agrupado de forma inequívoca, posiblemente debido a la escasez de especies raras clave como Bentheuphausia amblyops en krill y Amphionides reynaudii en Eucarida. Un estudio apoya la monofilia de Eucarida (con Mysida basal), [17] otro agrupa a Euphausiacea con Mysida (los Schizopoda), [12] mientras que otro más agrupa a Euphausiacea con Hoplocarida . [18]
Cronología
Ningún fósil existente puede asignarse inequívocamente a Euphausiacea. Se ha pensado que algunos taxones eumalacostráceos extintos eran eufausiáceos, como Anthracophausia , Crangopsis —ahora asignado a Aeschronectida (Hoplocarida) [8] —y Palaeomysis . [19] Todas las dataciones de los eventos de especiación se estimaron mediante métodos de reloj molecular , que ubicaron al último ancestro común de la familia del kril Euphausiidae (orden Euphausiacea menos Bentheuphausia amblyops ) como habiendo vivido en el Cretácico Inferior hace unos 130 millones de años . [12]
Las especies con distribuciones neríticas incluyen las cuatro especies del género Nyctiphanes . [22] Son muy abundantes a lo largo de las regiones de surgencia de los sistemas de corrientes de California , Humboldt , Benguela y Canarias . [23] [24] [25] Otra especie que solo tiene distribución nerítica es E. crystallorophias , que es endémica de la costa antártica. [26]
Las especies con distribuciones endémicas incluyen Nyctiphanes capensis , que se encuentra solo en la corriente de Benguela, [22] E. mucronata en la corriente de Humboldt, [27] y las seis especies de Euphausia nativas del Océano Austral.
En la Antártida, se conocen siete especies, [28] una en el género Thysanoessa ( T. macrura ) y seis en Euphausia . El krill antártico ( Euphausia superba ) vive comúnmente a profundidades que alcanzan los 100 m (330 pies), [29] mientras que el krill de hielo ( Euphausia crystallorophias ) alcanza una profundidad de 4.000 m (13.100 pies), aunque comúnmente habitan profundidades de como máximo 300-600 m (1.000-2.000 pies). [30] El krill realiza migraciones verticales diarias (DVM) en grandes enjambres, y los datos acústicos han demostrado que estas migraciones llegan hasta los 400 metros de profundidad. [31] Ambos se encuentran en latitudes al sur de 55° S , con E. crystallorophias dominando al sur de 74° S [32] y en regiones de hielo marino . Otras especies conocidas en el Océano Austral son E. frigida , E. longirostris , E. triacantha y E. vallentini . [33]
Anatomía y morfología
Explicación de la anatomía del krill utilizando Euphausia superba como modeloLas branquias del krill son visibles externamente.
Los krill son crustáceos y, como todos los crustáceos, tienen un exoesqueleto quitinoso . Tienen una anatomía similar a la de un decápodo estándar con sus cuerpos formados por tres partes : el cefalotórax está compuesto por la cabeza y el tórax , que están fusionados, y el abdomen , que lleva los diez apéndices nadadores, y el abanico de la cola . Esta capa exterior del krill es transparente en la mayoría de las especies.
El krill tiene ojos compuestos intrincados . Algunas especies se adaptan a diferentes condiciones de iluminación mediante el uso de pigmentos de protección . [34]
Tienen dos antenas y varios pares de patas torácicas llamadas pereiópodos o toracópodos , llamados así porque están unidos al tórax. Su número varía entre géneros y especies. Estas patas torácicas incluyen patas para alimentarse y patas para acicalarse.
El krill es probablemente el clado hermano de los decápodos porque todas las especies tienen cinco pares de patas nadadoras llamadas "nadadores" en común con estos últimos, muy similares a los de una langosta o un cangrejo de río de agua dulce .
A pesar de tener diez nadadores, también conocidos como pleópodos , el krill no puede considerarse decápodo. Carecen de patas terrestres verdaderas debido a que todos sus pereiópodos se han convertido en patas auxiliares de alimentación y acicalamiento. En Decapoda , hay diez pereiópodos funcionales , lo que les da su nombre; mientras que aquí no quedan pereiópodos locomotores . Tampoco hay diez pereiópodos de manera constante.
La mayoría de los krill miden entre 1 y 2 centímetros (0,4 y 0,8 pulgadas) de largo cuando son adultos. Unas pocas especies crecen hasta alcanzar tamaños del orden de 6 a 15 centímetros (2,4 a 5,9 pulgadas). La especie de krill más grande, Thysanopoda spinicaudata , vive en las profundidades del océano abierto . [35] El krill se puede distinguir fácilmente de otros crustáceos, como los camarones verdaderos , por sus branquias visibles externamente . [36]
A excepción de Bentheuphausia amblyops , los krill son animales bioluminiscentes que tienen órganos llamados fotóforos que pueden emitir luz. La luz se genera mediante una reacción de quimioluminiscencia catalizada por enzimas , en la que una luciferina (un tipo de pigmento) es activada por una enzima luciferasa . Los estudios indican que la luciferina de muchas especies de krill es un tetrapirrol fluorescente similar pero no idéntico a la luciferina de los dinoflagelados [37] y que es probable que el krill no produzca esta sustancia por sí mismo, sino que la adquiera como parte de su dieta, que contiene dinoflagelados. [38] Los fotóforos del krill son órganos complejos con lentes y capacidades de enfoque, y pueden rotarse mediante músculos. [39] La función precisa de estos órganos aún se desconoce; las posibilidades incluyen el apareamiento, la interacción social u orientación y como una forma de camuflaje contra la iluminación para compensar su sombra contra la luz ambiental superior. [40] [41]
Ecología
Procesos en la bomba biológica
El fitoplancton convierte el CO2 , que se ha disuelto de la atmósfera en los océanos superficiales (90 Gt año−1) en carbono orgánico particulado (POC) durante la producción primaria (~ 50 Gt C año−1). Luego, el fitoplancton es consumido por el krill y los pequeños herbívoros del zooplancton, que a su vez son depredados por niveles tróficos superiores. Cualquier fitoplancton no consumido forma agregados y, junto con los gránulos fecales del zooplancton, se hunde rápidamente y se exporta fuera de la capa mixta (< 12 Gt C año−1 14). El krill, el zooplancton y los microbios interceptan el fitoplancton en la superficie del océano y las partículas detríticas que se hunden en profundidad, consumiendo y respirando este POC a CO2 ( carbono inorgánico disuelto, DIC), de modo que solo una pequeña proporción del carbono producido en la superficie se hunde en el océano profundo (es decir, profundidades > 1000 m). A medida que el krill y el zooplancton más pequeño se alimentan, también fragmentan físicamente las partículas en trozos pequeños, que se hunden más lentamente o que no se hunden (a través de una alimentación descuidada, coprorrexia si se fragmentan las heces), lo que retarda la exportación de POC. Esto libera carbono orgánico disuelto (COD) ya sea directamente de las células o indirectamente a través de la solubilización bacteriana (círculo amarillo alrededor del COD). Las bacterias pueden luego remineralizar el COD a CID (CO2 , jardinería microbiana). El krill, el zooplancton más pequeño y los peces que migran verticalmente durante el día pueden transportar activamente carbono a la profundidad al consumir POC en la capa superficial durante la noche y metabolizarlo en sus profundidades de residencia mesopelágicas diurnas. Dependiendo del ciclo de vida de las especies, el transporte activo también puede ocurrir de forma estacional. Los números dados son flujos de carbono (Gt C año−1) en cuadros blancos y masas de carbono (Gt C) en cuadros oscuros. [42]
Alimentación
Muchos krill se alimentan por filtración : [24] sus apéndices más delanteros , los toracópodos, forman peines muy finos con los que pueden filtrar su alimento del agua. Estos filtros pueden ser muy finos en especies (como Euphausia spp.) que se alimentan principalmente de fitoplancton , en particular de diatomeas , que son algas unicelulares . El krill es en su mayoría omnívoro , [43] aunque unas pocas especies son carnívoras y se alimentan de zooplancton pequeño y larvas de peces . [44]
El krill es un elemento importante de la cadena alimentaria acuática . El krill convierte la producción primaria de sus presas en una forma adecuada para el consumo de animales más grandes que no pueden alimentarse directamente de las minúsculas algas. El krill del norte y algunas otras especies tienen una canasta filtrante relativamente pequeña y cazan activamente copépodos y zooplancton más grande. [44]
Depredación
Muchos animales se alimentan de krill, desde animales más pequeños como peces o pingüinos hasta otros más grandes como focas y ballenas barbadas . [45]
Las perturbaciones de un ecosistema que provocan una disminución de la población de krill pueden tener efectos de largo alcance. Por ejemplo, durante una floración de cocolitóforos en el mar de Bering en 1998 [46] , la concentración de diatomeas disminuyó en la zona afectada. El krill no puede alimentarse de los cocolitóforos más pequeños y, en consecuencia, la población de krill (principalmente E. pacifica ) en esa región disminuyó drásticamente. Esto, a su vez, afectó a otras especies: la población de pardelas disminuyó. Se cree que el incidente fue una de las razones por las que el salmón no desovaba esa temporada [47] .
Varios ciliados endoparasitoides unicelulares del género Collinia pueden infectar especies de krill y devastar las poblaciones afectadas. Se han descrito enfermedades de este tipo en Thysanoessa inermis en el mar de Bering y también en E. pacifica , Thysanoessa spinifera y T. gregaria frente a la costa del Pacífico de América del Norte. [48] [49] Algunos ectoparásitos de la familia Dajidae ( isópodos epicáridos ) afectan al krill (y también a los camarones y a los mísidos ); uno de estos parásitos es Oculophryxus bicaulis , que se encontró en el krill Stylocheiron affine y S. longicorne . Se adhiere al pedúnculo ocular del animal y succiona la sangre de su cabeza; aparentemente inhibe la reproducción del huésped, ya que ninguno de los animales afectados alcanzó la madurez. [50]
Las investigaciones preliminares indican que el krill puede digerir microplásticos de menos de 5 mm (0,20 pulgadas) de diámetro, descomponiéndolos y excretándolos nuevamente al medio ambiente en forma más pequeña. [52]
El ciclo de vida del krill se entiende relativamente bien, a pesar de pequeñas variaciones en los detalles de una especie a otra. [15] [24] Después de la eclosión del krill, pasan por varias etapas larvarias: nauplio , pseudometanauplio , metanauplio , caliptopsis y furcilia , cada una de las cuales se divide en subetapas. La etapa pseudometanauplio es exclusiva de las especies que ponen sus huevos dentro de un saco ovígero: los llamados "desovadores en saco". Las larvas crecen y mudan repetidamente a medida que se desarrollan, reemplazando su exoesqueleto rígido cuando se vuelve demasiado pequeño. Los animales más pequeños mudan con más frecuencia que los más grandes. Las reservas de vitelo dentro de su cuerpo nutren a las larvas durante la etapa de metanauplio.
En las etapas de calyptopsis, la diferenciación ha progresado lo suficiente como para que desarrollen una boca y un tracto digestivo, y comiencen a comer fitoplancton. Para entonces, sus reservas de vitelo se han agotado y las larvas deben haber alcanzado la zona fótica , las capas superiores del océano donde florecen las algas. Durante las etapas de furcilia, se agregan segmentos con pares de nadadores, comenzando por los segmentos más delanteros. Cada nuevo par se vuelve funcional solo en la siguiente muda. El número de segmentos agregados durante cualquiera de las etapas de furcilia puede variar incluso dentro de una especie dependiendo de las condiciones ambientales. [53] Después de la etapa final de furcilia, emerge un juvenil inmaduro con una forma similar a un adulto, y posteriormente desarrolla gónadas y madura sexualmente. [54]
Reproducción
La cabeza de una hembra de krill de la especie Nematoscelis difficilis, que desova en sacos , con su saco de cría. Los huevos tienen un diámetro de 0,3 a 0,4 milímetros (0,012 a 0,016 pulgadas)
Durante la época de apareamiento, que varía según la especie y el clima, el macho deposita un saco de esperma en la abertura genital de la hembra (llamado télico ). Las hembras pueden llevar varios miles de huevos en su ovario , que pueden representar hasta un tercio de la masa corporal del animal. [55] El krill puede tener múltiples crías en una temporada, con intervalos entre crías que duran del orden de días. [25] [56]
El krill emplea dos tipos de mecanismos de desove. [25] Las 57 especies de los géneros Bentheuphausia , Euphausia , Meganyctiphanes , Thysanoessa y Thysanopoda son "desovadores por difusión": la hembra libera los huevos fertilizados en el agua, donde generalmente se hunden, se dispersan y se quedan solos. Estas especies generalmente eclosionan en la etapa de nauplio 1, pero recientemente se ha descubierto que a veces eclosionan como metanauplio o incluso como etapas de calyptopis. [57] Las 29 especies restantes de los otros géneros son "desovadores en saco", donde la hembra lleva los huevos con ella, unidos a los pares más traseros de toracópodos hasta que eclosionan como metanauplios, aunque algunas especies como Nematoscelis difficilis pueden eclosionar como nauplios o pseudometanauplios. [58]
Muda
La muda se produce cuando un espécimen supera su exoesqueleto rígido. Los animales jóvenes, que crecen más rápido, mudan con más frecuencia que los mayores y más grandes. La frecuencia de la muda varía ampliamente según la especie y, incluso dentro de una misma especie, está sujeta a muchos factores externos como la latitud, la temperatura del agua y la disponibilidad de alimentos. La especie subtropical Nyctiphanes simplex , por ejemplo, tiene un período general entre mudas de dos a siete días: las larvas mudan en promedio cada cuatro días, mientras que los juveniles y los adultos lo hacen, en promedio, cada seis días. Para E. superba en el mar Antártico, se han observado períodos entre muda que varían entre 9 y 28 días dependiendo de la temperatura entre −1 y 4 °C (30 y 39 °F), y para Meganyctiphanes norvegica en el Mar del Norte los períodos entre muda también varían entre 9 y 28 días pero a temperaturas entre 2,5 y 15 °C (36,5 y 59,0 °F). [59] E. superba es capaz de reducir el tamaño de su cuerpo cuando no hay suficiente alimento disponible, mudando también cuando su exoesqueleto se vuelve demasiado grande. [60] También se ha observado una contracción similar en E. pacifica , una especie que se encuentra en el Océano Pacífico desde zonas polares a templadas, como una adaptación a temperaturas del agua anormalmente altas. También se ha postulado la contracción para otras especies de krill de zonas templadas. [61]
Esperanza de vida
Algunas especies de krill de altas latitudes pueden vivir más de seis años (por ejemplo, Euphausia superba ); otras, como la especie de latitudes medias Euphausia pacifica , viven solo dos años. [7] La longevidad de las especies subtropicales o tropicales es aún más corta, por ejemplo, Nyctiphanes simplex , que generalmente vive solo de seis a ocho meses. [62]
Enjambre
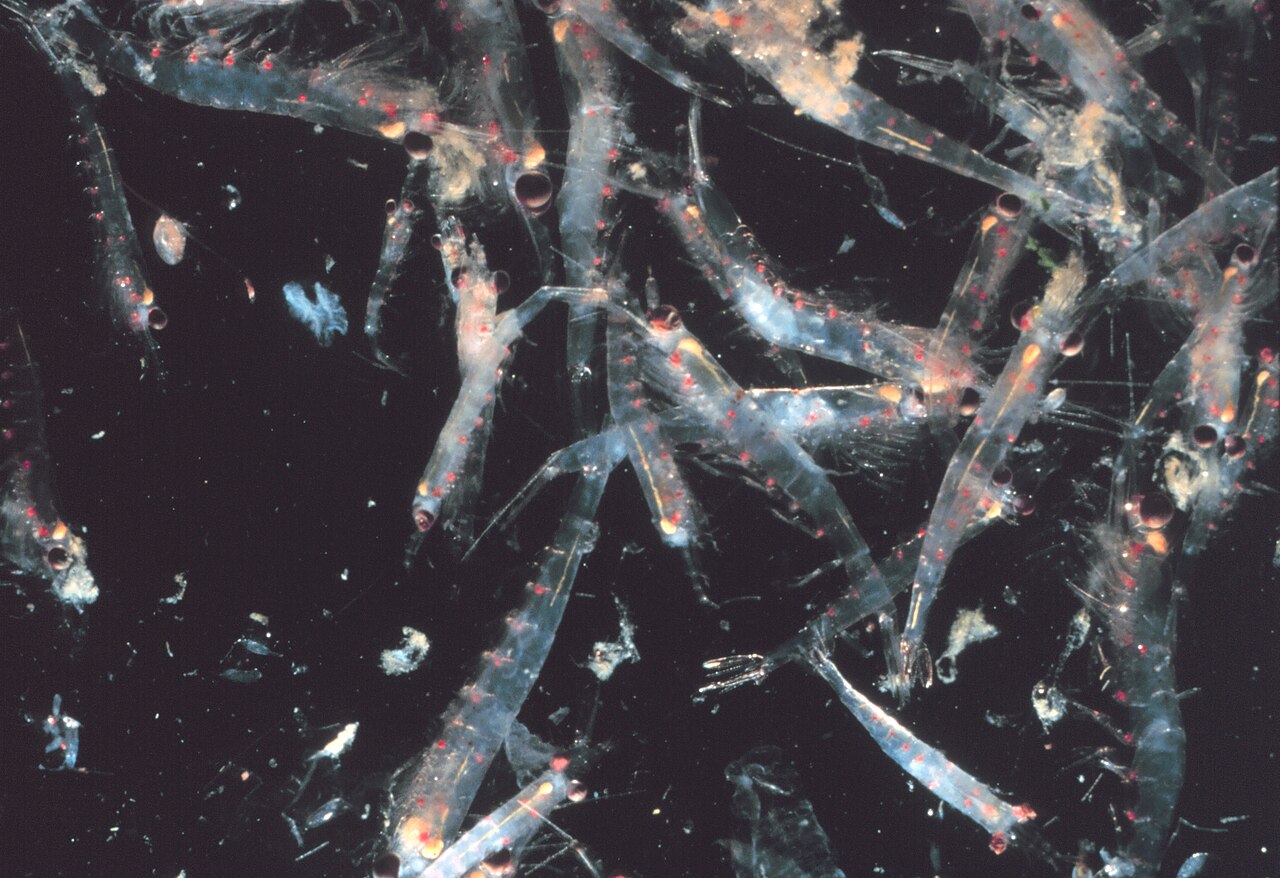
Un enjambre de krill
La mayoría de los krill son animales que forman enjambres ; los tamaños y densidades de estos enjambres varían según la especie y la región. En el caso de Euphausia superba , los enjambres alcanzan entre 10.000 y 60.000 individuos por metro cúbico. [63] [64] La formación de enjambres es un mecanismo defensivo que confunde a los depredadores más pequeños que quisieran localizar a los individuos. En 2012, Gandomi y Alavi presentaron lo que parece ser un algoritmo estocástico exitoso para modelar el comportamiento de los enjambres de krill. El algoritmo se basa en tres factores principales: " (i) movimiento inducido por la presencia de otros individuos (ii) actividad de alimentación y (iii) difusión aleatoria". [65]
El krill suele seguir una migración vertical diurna . Se ha asumido que pasan el día a mayores profundidades y ascienden durante la noche hacia la superficie. Cuanto más profundo descienden, más reducen su actividad, [66] aparentemente para reducir los encuentros con depredadores y conservar energía. La actividad natatoria del krill varía con la saciedad del estómago. Los animales saciados que se han estado alimentando en la superficie nadan menos activamente y, por lo tanto, se hunden por debajo de la capa mixta. [67] A medida que se hunden, producen heces que desempeñan un papel en el ciclo del carbono antártico . El krill con el estómago vacío nada más activamente y, por lo tanto, se dirige hacia la superficie.
La migración vertical puede ocurrir 2 o 3 veces al día. Algunas especies (por ejemplo, Euphausia superba , E. pacifica , E. hanseni , Pseudeuphausia latifrons y Thysanoessa spinifera ) forman enjambres superficiales durante el día con fines de alimentación y reproducción, aunque este comportamiento es peligroso porque las hace extremadamente vulnerables a los depredadores. [68]
Estudios experimentales que utilizan Artemia salina como modelo sugieren que las migraciones verticales de krill de varios cientos de metros, en grupos de decenas de metros de profundidad, podrían crear colectivamente suficientes chorros de agua descendentes para tener un efecto significativo en la mezcla oceánica. [69]
Los enjambres densos pueden provocar un frenesí alimentario entre peces, aves y mamíferos depredadores, especialmente cerca de la superficie. Cuando se los molesta, los enjambres se dispersan y se ha observado que algunos individuos mudan de piel al instante, dejando atrás la exuvia como señuelo. [70]
El krill normalmente nada a un ritmo de 5-10 cm/s (2-3 longitudes corporales por segundo), [71] utilizando sus nadadores para propulsarse. Sus migraciones más grandes están sujetas a las corrientes oceánicas. Cuando están en peligro, muestran una reacción de escape llamada lobstering : moviendo sus estructuras caudales , el telson y los urópodos , se mueven hacia atrás a través del agua con relativa rapidez, alcanzando velocidades en el rango de 10 a 27 longitudes corporales por segundo, lo que para krill grandes como E. superba significa alrededor de 0,8 m/s (3 pies/s). [72] Su rendimiento de natación ha llevado a muchos investigadores a clasificar al krill adulto como formas de vida micronectónicas , es decir, pequeños animales capaces de moverse individualmente contra corrientes (débiles). Las formas larvarias de krill generalmente se consideran zooplancton. [73]
Ciclos biogeoquímicos
El papel del krill antártico en los ciclos biogeoquímicos
El krill (en forma de enjambres e individuos) se alimenta de fitoplancton en la superficie (1), dejando solo una proporción que se hunda en forma de agregados fitodetríticos (2), que se desintegran fácilmente y pueden no hundirse por debajo de la termoclina permanente. El krill también libera heces fecales (3) mientras se alimenta, que pueden hundirse en las profundidades del mar, pero pueden ser consumidas (coprofagia) y degradadas a medida que descienden (4) por el krill, las bacterias y el zooplancton. En la zona de hielo marginal, el flujo de heces fecales puede alcanzar mayores profundidades (5). El krill también libera mudas, que se hunden y contribuyen al flujo de carbono (6). El krill libera nutrientes durante la alimentación descuidada, la excreción y la egestión, como el hierro y el amonio (7, véase la figura 2 para otros nutrientes liberados), y si se liberan cerca de la superficie pueden estimular la producción de fitoplancton y una mayor reducción del CO2 atmosférico . Algunos krill adultos residen permanentemente en las profundidades de la columna de agua, consumiendo material orgánico en profundidad (8). Todo el carbono (como materia orgánica o como CO2 ) que se hunda por debajo de la termoclina permanente se elimina de la mezcla estacional y permanecerá almacenado en las profundidades del océano durante al menos un año (9). Los movimientos de natación del krill adulto que migra puede mezclar agua rica en nutrientes de las profundidades (10), estimulando aún más la producción primaria. Otros krill adultos se alimentan en el fondo marino, liberando el CO2 respirado en profundidad y pueden ser consumidos por depredadores demersales (11). El krill larvario, que en el océano Austral reside bajo el hielo marino, experimenta una extensa migración vertical diurna (12), transfiriendo potencialmente CO2 por debajo de la termoclina permanente. El krill es consumido por muchos depredadores, incluidas las ballenas barbadas (13), lo que lleva al almacenamiento de parte del carbono del krill como biomasa durante décadas antes de que la ballena muera, se hunda en el fondo marino y sea consumida por organismos de aguas profundas. [42]
Ciclo de nutrientes por parte de un krill individual
Cuando el krill muda, libera calcio, flúor y fósforo disueltos del exoesqueleto (1). La quitina (material orgánico) que forma el exoesqueleto contribuye al flujo de partículas orgánicas que se hunden en las profundidades del océano. El krill respira una parte de la energía derivada del consumo de fitoplancton u otros animales en forma de dióxido de carbono (2); cuando nada desde aguas medias/profundas hasta la superficie en grandes cardúmenes, el krill mezcla agua, lo que potencialmente lleva nutrientes a las aguas superficiales pobres en nutrientes (3); el amonio y el fosfato se liberan de las branquias y cuando excretan, junto con el carbono orgánico disuelto, nitrógeno (p. ej., urea) y fósforo (COD, DON y DOP, 2 y 4). El krill libera gránulos fecales que se hunden rápidamente y que contienen carbono orgánico particulado, nitrógeno y fósforo (POC, PON y POP) y hierro, el último de los cuales es biodisponible cuando se lixivia a las aguas circundantes junto con el COD, el DON y el DOP (5). [42]
Usos humanos
Placas de krill antártico ultracongeladas para su uso como alimento animal y materia prima para cocinar
Historia de la cosecha
El krill se ha cosechado como fuente de alimento para humanos y animales domésticos desde al menos el siglo XIX, y posiblemente antes en Japón, donde se lo conocía como okiami . La pesca a gran escala se desarrolló a fines de la década de 1960 y principios de la de 1970, y ahora se realiza solo en aguas antárticas y en los mares que rodean Japón. Históricamente, las naciones pesqueras de krill más grandes fueron Japón y la Unión Soviética o, después de la disolución de esta última, Rusia y Ucrania . [77] La cosecha máxima, que en 1983 fue de aproximadamente 528.000 toneladas solo en el Océano Austral (de las cuales la Unión Soviética capturó el 93%), ahora se gestiona como precaución contra la sobrepesca. [78]
La captura anual antártica se estabilizó en alrededor de 100.000 toneladas, lo que representa aproximadamente una quincuagésima parte de la cuota de captura de la CCRVMA. [80] El principal factor limitante fueron probablemente los altos costos junto con cuestiones políticas y legales. [81] La pesquería japonesa se saturó en unas 70.000 toneladas. [82]
Aunque el krill se encuentra en todo el mundo, se prefiere la pesca en los océanos australes porque es más "capturable" y abundante en esas regiones. En particular, en los mares antárticos, que se consideran prístinos, se considera un "producto limpio". [77]
En 2018 se anunció que casi todas las empresas pesqueras de krill que operan en la Antártida abandonarán sus operaciones en grandes áreas alrededor de la Península Antártica a partir de 2020, incluidas las "zonas de amortiguación" alrededor de las colonias de reproducción de pingüinos. [83]
Aunque la biomasa total de krill antártico puede ser tan abundante como 400 millones de toneladas, el impacto humano sobre esta especie clave está creciendo, con un aumento del 39% en el rendimiento pesquero total a 294.000 toneladas durante 2010-2014. [80] Los principales países involucrados en la recolección de krill son Noruega (56% de la captura total en 2014), la República de Corea (19%) y China (18%). [80]
El krill es una fuente rica de proteínas y ácidos grasos omega-3 que se está desarrollando a principios del siglo XXI como alimento humano, suplementos dietéticos como cápsulas de aceite, alimento para ganado y alimento para mascotas . [77] [79] [84] El krill tiene un sabor salado con un sabor a pescado algo más fuerte que el camarón. Para el consumo masivo y los productos preparados comercialmente, deben pelarse para eliminar el exoesqueleto no comestible . [84]
El krill (y otros camarones planctónicos , en particular Acetes spp.) se consumen más ampliamente en el sudeste asiático, donde se fermentan (con las cáscaras intactas) y generalmente se muelen finamente para hacer pasta de camarones . Se puede saltear y comer acompañado de arroz blanco o usarse para agregar sabores umami a una amplia variedad de platos tradicionales. [86] [87] El líquido del proceso de fermentación también se cosecha como salsa de pescado . [88]
Robótica bioinspirada
Los krill son nadadores ágiles en el régimen de número de Reynolds intermedio , en el que no hay muchas soluciones para la robótica submarina no tripulada, y han inspirado plataformas robóticas tanto para estudiar su locomoción como para encontrar soluciones de diseño para robots submarinos. [89]
^ "Eufáusidos (krill)". Gobierno de Canadá . Ministerio de Pesca y Océanos de Canadá. 6 de abril de 2022. Consultado el 18 de abril de 2024. En las costas este y oeste de Canadá se encuentran muchas especies diferentes de eufáusidos.
^ Crustáceos: Euphasiacea - Oxford Academic
^ "Krill". Diccionario Etimológico Online . Consultado el 22 de junio de 2010 .
^ A. Atkinson; V. Siegel; EA Pakhomov; MJ Jessopp; V. Loeb (2009). "Una reevaluación de la biomasa total y la producción anual de krill antártico" (PDF) . Investigación en aguas profundas, parte I . 56 (5): 727–740. Bibcode :2009DSRI...56..727A. doi :10.1016/j.dsr.2008.12.007.
↑ Siegel V (2011). Siegel V (ed.). "Euphausiidae Dana, 1852". Base de datos mundial de eufausiacea . Registro mundial de especies marinas . Consultado el 25 de noviembre de 2011 .
^ E. Brinton (1962). "La distribución de los eufáusidos del Pacífico". Bull. Scripps Inst. Oceanogr . 8 (2): 51–270.
^ ab S. Nicol; Y. Endo (1999). "Pesquerías de krill: desarrollo, gestión e implicaciones para el ecosistema". Recursos vivos acuáticos . 12 (2): 105–120. doi :10.1016/S0990-7440(99)80020-5. S2CID 84158071.
^ abcd Andreas Maas; Dieter Waloszek (2001). "Desarrollo larvario de Euphausia superba Dana, 1852 y un análisis filogenético de las Euphausiacea" (PDF) . Hydrobiologia . 448 : 143–169. doi :10.1023/A:1017549321961. S2CID 32997380. Archivado desde el original (PDF) el 18 de julio de 2011.
^ Bernadette Casanova (1984). "Phylogénie des Euphausiacés (Crustacés Eucarides)" [Filogenia de las Euphausiacea (Crustacea: Eucarida)]. Bulletin du Muséum National d'Histoire Naturelle (en francés). 4 : 1077–1089.
^ abc Bernadette Casanova (2003). "Ordre des Euphausiacea Dana, 1852". Crustáceos . 76 (9): 1083-1121. doi :10.1163/156854003322753439. JSTOR 20105650.
^ M. Eugenia D'Amato; Gordon W. Harkins; Tulio de Oliveira; Peter R. Teske; Mark J. Gibbons (2008). «Molecular dating and biogeography of the neritic krill Nyctiphanes» (PDF) . Biología Marina . 155 (2): 243–247. doi :10.1007/s00227-008-1005-0. S2CID 17750015. Archivado desde el original (PDF) el 17 de marzo de 2012 . Consultado el 4 de julio de 2010 .
^ abc Simon N. Jarman (2001). "La historia evolutiva del krill inferida a partir del análisis de la secuencia de la subunidad grande nuclear del ADNr". Revista biológica de la Sociedad Linneana . 73 (2): 199–212. doi : 10.1111/j.1095-8312.2001.tb01357.x .
^ Xin Shen; Haiqing Wang; Minxiao Wang; Bin Liu (2011). "La secuencia completa del genoma mitocondrial de Euphausia pacifica (Malacostraca: Euphausiacea) revela un nuevo orden genético y repeticiones en tándem inusuales". Genoma . 54 (11): 911–922. doi :10.1139/g11-053. PMID 22017501.
^ Johan Erik Vesti Boas (1883). "Studien über die Verwandtschaftsbeziehungen der Malacostraken" [Estudios sobre las relaciones de los Malacostraca]. Morphologisches Jahrbuch (en alemán). 8 : 485–579.
^ Isabella Gordon (1955). "Posición sistemática de las Euphausiacea". Nature . 176 (4489): 934. Bibcode :1955Natur.176..934G. doi : 10.1038/176934a0 . S2CID 4225121.
^ Trisha Spears, Ronald W. DeBry, Lawrence G. Abele y Katarzyna Chodyl (2005). Boyko, Christopher B. (ed.). "Monofilia de peracáridos y filogenia interordinal inferida a partir de secuencias de ADN ribosómico de subunidades pequeñas nucleares (Crustacea: Malacostraca: Peracarida)" (PDF) . Actas de la Sociedad Biológica de Washington . 118 (1): 117–157. doi :10.2988/0006-324X(2005)118[117:PMAIPI]2.0.CO;2. S2CID 85557065.{{cite journal}}: CS1 maint: varios nombres: lista de autores ( enlace )
^ K. Meland; E. Willassen (2007). "La desunión de" Mysidacea "(Crustacea)". Filogenética molecular y evolución . 44 (3): 1083-1104. CiteSeerX 10.1.1.653.5935 . doi :10.1016/j.ympev.2007.02.009. PMID 17398121.
^ ab D'Amato, ME et al. : "Datación molecular y biogeografía del krill nerítico Nyctiphanes Archivado el 17 de marzo de 2012 en Wayback Machine ", en Marine Biology vol. 155, no. 2 , pp. 243–247, agosto de 2008.
^ Volker Siegel (2011). V. Siegel (ed.). "Nyctiphanes Sars, 1883". Base de datos mundial de eufausias . Registro mundial de especies marinas . Consultado el 18 de junio de 2011 .
^ abc J. Mauchline; LR Fisher (1969). La biología de los eufáusidos . Avances en biología marina. Vol. 7. Academic Press . ISBN.978-7-7708-3615-2.
^ abc Jaime Gómez-Gutiérrez; Carlos J. Robinson (2005). "Tiempo de desarrollo embrionario, larval temprano, mecanismo de eclosión y período de intercría del eufáusido de desove en saco Nyctiphanes simplex Hansen". Journal of Plankton Research . 27 (3): 279–295. doi : 10.1093/plankt/fbi003 .
^ SN Jarman; NG Elliott; S. Nicol; A. McMinn (2002). "Diferenciación genética en el krill costero antártico Euphausia crystallorophias". Heredity . 88 (4): 280–287. doi : 10.1038/sj.hdy.6800041 . PMID 11920136.
^ R. Escribano; V. Marín; C. Irribarren (2000). "Distribución de Euphausia mucronata en la zona de surgencia de Península Mejillones, norte de Chile: la influencia de la capa mínima de oxígeno". Marina científica . 64 (1): 69–77. doi : 10.3989/scimar.2000.64n169 .
^ P. Brueggeman. "Euphausia crystallorophias". Guía de campo submarina de la isla Ross y el estrecho McMurdo, Antártida . Universidad de California, San Diego .
^ "Krill, Euphausia superba". MarineBio.org . Consultado el 25 de febrero de 2009 .
^ JA Kirkwood (1984). "Una guía sobre las eufausiáceas del océano Austral". Notas de investigación de ANARE . 1 : 1–45.
^ Bianchi, Daniele; Mislan, KAS (enero de 2016). "Patrones globales de tiempos y velocidades de migración vertical diaria a partir de datos acústicos". Limnología y Oceanografía . 61 (1). doi : 10.1002/lno.10219 .
^ A. Sala; M. Azzali; A. Russo (2002). "Krill del mar de Ross: distribución, abundancia y demografía de Euphausia superba y Euphausia crystallorophias durante la expedición antártica italiana (enero-febrero de 2000)". Scientia Marina . 66 (2): 123–133. doi : 10.3989/scimar.2002.66n2123 .
^ GW Hosie; M. Fukuchi; S. Kawaguchi (2003). "Desarrollo del estudio del registrador continuo de plancton del océano Austral" (PDF) . Progreso en Oceanografía . 58 (2–4): 263–283. Bibcode :2003PrOce..58..263H. doi :10.1016/j.pocean.2003.08.007.[ enlace muerto permanente ]
^ E. Gaten. "Meganyctiphanes norvegica". Universidad de Leicester . Archivado desde el original el 1 de julio de 2009. Consultado el 25 de febrero de 2009 .
^ E. Brinton (1953). " Thysanopoda spinicauda , un nuevo crustáceo eufáusido gigante batipelágico, con notas comparativas sobre T. cornuta y T. egregia ". Revista de la Academia de Ciencias de Washington . 43 : 408–412.
^ "Euphausiacea". Instituto de Acuicultura y Pesca de Tasmania. Archivado desde el original el 30 de septiembre de 2009. Consultado el 6 de junio de 2010 .
^ O. Shimomura (1995). "Los roles de los dos componentes altamente inestables F y P involucrados en la bioluminiscencia de los camarones eufáusidos". Journal of Bioluminescence and Chemiluminescence . 10 (2): 91–101. doi :10.1002/bio.1170100205. PMID 7676855.
^ JC Dunlap; JW Hastings; O. Shimomura (1980). "Reactividad cruzada entre los sistemas emisores de luz de organismos distantemente relacionados: nuevo tipo de compuesto emisor de luz". Actas de la Academia Nacional de Ciencias . 77 (3): 1394–1397. Bibcode :1980PNAS...77.1394D. doi : 10.1073/pnas.77.3.1394 . JSTOR 8463. PMC 348501 . PMID 16592787.
^ PJ Herring; EA Widder (2001). "Bioluminiscencia en el plancton y el necton". En JH Steele; SA Thorpe; KK Turekian (eds.). Enciclopedia de la ciencia oceánica . Vol. 1. Academic Press , San Diego. págs. 308–317. ISBN978-0-12-227430-5.
^ SM Lindsay; MI Latz (1999). Evidencia experimental de contrasombreado luminiscente por algunos crustáceos eufáusidos . Reunión de Ciencias Acuáticas de la Sociedad Estadounidense de Limnología y Oceanografía (ASLO). Santa Fe.
^ Sönke Johnsen (2005). "El rojo y el negro: bioluminiscencia y el color de los animales en las profundidades marinas" (PDF) . Integrative and Comparative Biology . 4 (2): 234–246. doi : 10.1093/icb/45.2.234 . PMID 21676767. S2CID 247718. Archivado desde el original (PDF) el 2 de octubre de 2005.
^ abcd Cavan, EL, Belcher, A., Atkinson, A., Hill, SL, Kawaguchi, S., McCormack, S., Meyer, B., Nicol, S., Ratnarajah, L., Schmidt, K. y Steinberg, DK (2019) "La importancia del krill antártico en los ciclos biogeoquímicos". Nature communications , 10 (1): 1–13. doi :10.1038/s41467-019-12668-7.El material fue copiado de esta fuente, que está disponible bajo una Licencia Creative Commons Atribución 4.0 Internacional.
^ GC Cripps; A. Atkinson (2000). "Composición de ácidos grasos como indicador de carnivoría en el krill antártico, Euphausia superba ". Revista Canadiense de Ciencias Pesqueras y Acuáticas . 57 (S3): 31–37. doi :10.1139/f00-167.
^ ab Olav Saether; Trond Erling Ellingsen; Viggo Mohr (1986). "Lípidos de krill del Atlántico norte" (PDF) . Revista de investigación de lípidos . 27 (3): 274–285. PMID 3734626.
^ MJ Schramm (10 de octubre de 2007). «Tiny Krill: Giants in Marine Food Chain» (Krill diminuto: gigantes en la cadena alimentaria marina). Programa Nacional de Santuarios Marinos de la NOAA . Consultado el 4 de junio de 2010 .
^ J. Weier (1999). "Las corrientes cambiantes colorean el mar de Bering con un nuevo tono de azul". Observatorio de la Tierra de la NOAA . Consultado el 15 de junio de 2005 .
^ RD Brodeur; GH Kruse; PA Livingston; G. Walters; J. Ianelli; GL Swartzman; M. Stepanenko; T. Wyllie-Echeverria (1998). Informe preliminar del taller internacional de FOCI sobre las condiciones recientes en el mar de Bering . NOAA . págs. 22–26.
^ J. Roach (17 de julio de 2003). «Los científicos descubren un misterioso asesino del krill». National Geographic News . Archivado desde el original el 24 de julio de 2003.
^ J. Gómez-Gutiérrez; WT Peterson; A. de Robertis; RD Brodeur (2003). "Mortalidad masiva de krill causada por ciliados parasitoides". Science . 301 (5631): 339. doi :10.1126/science.1085164. PMID 12869754. S2CID 28471713.
^ JD Shields; J. Gómez-Gutiérrez (1996). " Oculophryxus bicaulis , un nuevo género y especie de isópodo dajid parásito del eufáusido Stylocheiron affine Hansen". Revista Internacional de Parasitología . 26 (3): 261–268. doi :10.1016/0020-7519(95)00126-3. PMID 8786215.
^ Rusty Dornin (6 de julio de 1997). «Disminuyen las poblaciones de krill antártico». CNN . Consultado el 18 de junio de 2011 .
^ Dawson, Amanda L; Kawaguchi, So; King, Catherine K; Townsend, Kathy A; King, Robert; Huston, Wilhelmina M; Bengtson Nash, Susan M (2018). "Convertir microplásticos en nanoplásticos mediante fragmentación digestiva por krill antártico". Nature Communications . 9 (1): 1001. Bibcode :2018NatCo...9.1001D. doi :10.1038/s41467-018-03465-9. PMC 5843626 . PMID 29520086.
^ MD Knight (1984). "Variación en la morfogénesis larval dentro de la población de Euphausia pacifica de la bahía del sur de California desde el invierno hasta el verano, 1977-1978" (PDF) . Informe CalCOFI . XXV . Archivado desde el original (PDF) el 3 de agosto de 2019. Consultado el 5 de noviembre de 2017 .
^ RM Ross; LB Quetin (1986). "¿Cuán productivo es el krill antártico?". BioScience . 36 (4): 264–269. doi :10.2307/1310217. JSTOR 1310217.
^ Janine Cuzin-Roudy (2000). "Reproducción estacional, desove múltiple y fecundidad en el krill del norte, Meganyctiphanes norvegica , y el krill antártico, Euphausia superba ". Revista Canadiense de Ciencias Pesqueras y Acuáticas . 57 (S3): 6–15. doi :10.1139/f00-165.
^ J. Gómez-Gutiérrez (2002). "Mecanismo de eclosión y eclosión retardada de los huevos de tres especies de eufáusidos que desovan al voleo en condiciones de laboratorio". Journal of Plankton Research . 24 (12): 1265–1276. doi : 10.1093/plankt/24.12.1265 .
^ E. Brinton; MD Ohman; AW Townsend; MD Caballero; AL Bridgeman (2000). Eufaúsidos del océano mundial. Serie de CD-ROM de la base de datos mundial sobre biodiversidad, Springer Verlag . ISBN978-3-540-14673-5Archivado desde el original el 26 de febrero de 2012 . Consultado el 4 de diciembre de 2009 .
^ F. Buchholz (2003). "Experimentos sobre la fisiología del krill del sur y del norte, Euphausia superba y Meganyctiphanes norvegica , con énfasis en la muda y el crecimiento: una revisión". Comportamiento y fisiología marina y de agua dulce . 36 (4): 229–247. Bibcode :2003MFBP...36..229B. doi :10.1080/10236240310001623376. S2CID 85121989.
^ H.-C. Shin; S. Nicol (2002). "Uso de la relación entre el diámetro ocular y la longitud corporal para detectar los efectos de la inanición a largo plazo en el krill antártico Euphausia superba". Marine Ecology Progress Series . 239 : 157–167. Bibcode :2002MEPS..239..157S. doi : 10.3354/meps239157 .
^ B. Marinovic; M. Mangel (1999). "El krill puede encogerse como una adaptación ecológica a entornos temporalmente desfavorables" (PDF) . Ecology Letters . 2 : 338–343.
^ JG Gómez (1995). "Patrones de distribución, abundancia y dinámica poblacional de los eufáusidos Nyctiphanes simplex y Euphausia eximia frente a la costa oeste de Baja California, México" (PDF) . Marine Ecology Progress Series . 119 : 63–76. Bibcode :1995MEPS..119...63G. doi : 10.3354/meps119063 .
^ U. Kils; P. Marshall (1995). " Der Krill, wie er schwimmt und frisst - neue Einsichten mit neuen Methoden (" El krill antártico - cómo nada y se alimenta - nuevos conocimientos con nuevos métodos ")". En I. Hempel; G. Hempel (eds.). Biologie der Polarmeere – Erlebnisse und Ergebnisse ( Experiencias y resultados de la biología de los océanos polares ) . Fischer Verlag . págs. 201–210. ISBN978-3-334-60950-7.
^ R. Piper (2007). Animales extraordinarios: una enciclopedia de animales curiosos e inusuales . Greenwood Press . ISBN978-0-313-33922-6.
^ Gandomi, AH; Alavi, AH (2012). "Krill Herd: Un nuevo algoritmo de optimización de inspiración biológica". Communications in Nonlinear Science and Numerical Simulation . 17 (12): 4831–4845. Bibcode :2012CNSNS..17.4831G. doi :10.1016/j.cnsns.2012.05.010.
^ JS Jaffe; MD Ohmann; A. de Robertis (1999). "Estimaciones de sonar de los niveles de actividad diurna de Euphausia pacifica en Saanich Inlet" (PDF) . Revista Canadiense de Pesca y Ciencias Acuáticas . 56 (11): 2000–2010. doi :10.1139/cjfas-56-11-2000. S2CID 228567512. Archivado desde el original (PDF) el 20 de julio de 2011.
^ Geraint A. Tarling; Magnus L. Johnson (2006). "La saciedad le da al krill esa sensación de hundimiento". Current Biology . 16 (3): 83–84. doi : 10.1016/j.cub.2006.01.044 . PMID 16461267.
^ Dan Howard (2001). "Krill" (PDF) . En Herman A. Karl; John L. Chin; Edward Ueber; Peter H. Stauffer; James W. Hendley II (eds.). Más allá del Golden Gate: oceanografía, geología, biología y cuestiones ambientales en el golfo de los Farallones. Servicio Geológico de los Estados Unidos . págs. 133–140. Circular 1198. Consultado el 8 de octubre de 2011 .
^ Wishart, Skye (julio-agosto de 2018). "El efecto krill". New Zealand Geographic (152): 24.
^ David A. Demer; Stéphane G. Conti (2005). "Nuevo modelo de intensidad de objetivo indica más krill en el océano Austral". ICES Journal of Marine Science . 62 (1): 25–32. doi : 10.1016/j.icesjms.2004.07.027 .
^ U. Kils (1982). «Comportamiento natatorio, rendimiento natatorio y balance energético del krill antártico Euphausia superba». BIOMASS Scientific Series 3, BIOMASS Research Series : 1–122. Archivado desde el original el 2 de junio de 2020 . Consultado el 11 de noviembre de 2017 .
^ S. Nicol; Y. Endo (1997). "Pesquerías de krill en el mundo". Documento técnico de pesca de la FAO . 367 .
^ Ratnarajah, L., Bowie, AR, Lannuzel, D., Meiners, KM y Nicol, S. (2014) "El papel biogeoquímico de las ballenas barbadas y el krill en el ciclo de nutrientes del Océano Austral". PLOS ONE , 9 (12): e114067. doi :10.1371/journal.pone.0114067
^ Hopkins, TL, Ainley, DG, Torres, JJ, Lancraft, TM, 1993. Estructura trófica en aguas abiertas de la zona de hielo marginal en la región de la confluencia Scotia-Weddell durante la primavera (1983). Polar Biology 13, 389–397.
^ Lancraft, TM, Relsenbichler, KR, Robinson, BH, Hopkins, TL, Torres, JJ, 2004. Una comunidad de micronecton y macrozooplancton dominada por krill en el Pasaje de Croker, Antártida, con una estimación de la depredación por peces. Deep-Sea Research II 51, 2247–2260.
^ abc Grossman, Elizabeth (14 de julio de 2015). "Los científicos consideran si es necesario proteger al krill de la caza excesiva por parte de los humanos". Public Radio International (PRI) . Consultado el 1 de abril de 2017 .
^ "Pesquerías de krill y sostenibilidad: krill antártico (Euphausia superba)". Comisión para la Conservación de los Recursos Vivos Marinos Antárticos. 23 de abril de 2015. Consultado el 1 de abril de 2017 .
^ ab Schiermeier, Q (2010). "Los ecologistas temen una crisis del krill antártico". Nature . 467 (7311): 15. doi : 10.1038/467015a . PMID 20811427.
^ abc «Krill – biología, ecología y pesca». Comisión para la Conservación de los Recursos Vivos Marinos Antárticos. 28 de abril de 2015. Consultado el 1 de abril de 2017 .
^ Minturn J. Wright (1987). "La propiedad de la Antártida, sus recursos vivos y minerales". Revista de Derecho y Medio Ambiente . 4 (2): 49–78.
^ S. Nicol; J. Foster (2003). "Tendencias recientes en la pesca del krill antártico". Recursos vivos acuáticos . 16 : 42–45. doi :10.1016/S0990-7440(03)00004-4.
^ Josh, Gabbatiss (10 de julio de 2018). "La industria pesquera de krill respalda un santuario oceánico antártico masivo para proteger a pingüinos, focas y ballenas". The Independent . Consultado el 10 de julio de 2018 .
^ ab "¿Por qué el krill?". Southwest Fisheries Science Center, Administración Nacional Oceánica y Atmosférica de EE. UU. 22 de noviembre de 2016. Consultado el 1 de abril de 2017 .
^ Cheeseman MA (22 de julio de 2011). "Aceite de krill: Carta de respuesta de la agencia, Aviso GRAS n.º GRN 000371". FDA de EE. UU . . Consultado el 3 de junio de 2015 .
^ Omori, M. (1978). "Pesquerías de zooplancton del mundo: una revisión". Biología marina . 48 (3): 199–205. doi :10.1007/BF00397145. S2CID 86540101.
^ Pongsetkul, Jaksuma; Benjakul, Soottawat; Sampavapol, Punnanee; Osako, Kazufumi; Faithong, Nandhsha (17 de septiembre de 2014). "Composición química y propiedades físicas de la pasta de camarón salada (Kapi) producida en Tailandia". Investigación Acuática Internacional . 6 (3): 155–166. doi : 10.1007/s40071-014-0076-4 .
^ Abe, Kenji; Suzuki, Kenji; Hashimoto, Kanehisa (1979). "Utilización de krill como material para salsa de pescado". Nippon Suisan Gakkaishi . 45 (8): 1013–1017. doi : 10.2331/suisan.45.1013 .
^ Oliveira Santos, Sara; Ataque, Nils; Su, Yunxing; Cuenca-Jiménez, Francisco; Morales-López, Oscar; Gómez-Valdez, P. Antonio; M Wilhelmus, Monica (13 de junio de 2023). "Pleobot: una solución robótica modular para natación metacrónica". Informes científicos . 13 (1): 9574. Código bibliográfico : 2023NatSR..13.9574O. doi :10.1038/s41598-023-36185-2. PMC 10264458 . PMID 37311777.
Lectura adicional
Boden, Brian P.; Johnson, Martin W .; Brinton, Edward: "Euphausiacea (Crustacea) del Pacífico Norte". Boletín del Instituto Scripps de Oceanografía . Volumen 6, número 8, 1955.
Brinton, Edward : "Eufáusidos de las aguas del sudeste asiático". Naga Report, volumen 4, parte 5. La Jolla: Universidad de California, Scripps Institution of Oceanography, 1975.
Conway, DVP; White, RG; Hugues-Dit-Ciles, J.; Galienne, CP; Robins, DB: Guía del zooplancton costero y de superficie del océano Índico sudoccidental Archivado el 23 de octubre de 2012 en Wayback Machine , Orden Euphausiacea, Publicación ocasional de la Asociación de Biología Marina del Reino Unido No. 15, Plymouth, Reino Unido, 2003.
Everson, I. (ed.): Krill: biología, ecología y pesca . Oxford, Blackwell Science; 2000. ISBN 0-632-05565-0 .
Hamner, William M. (mayo de 1984). "Krill: ¿una fuente inexplotada de riqueza marina?". National Geographic . Vol. 165, núm. 5. págs. 626–642. ISSN 0027-9358. OCLC 643483454.
Mauchline, J.: Euphausiacea: Adultos Archivado el 15 de mayo de 2011 en Wayback Machine , Conseil International pour l'Exploration de la Mer, 1971. Fichas de identificación de krill adulto con numerosos dibujos lineales. Archivo PDF , 2 Mb .
Mauchline, J.: Euphausiacea: Larvas Archivado el 19 de abril de 2012 en Wayback Machine , Conseil International pour l'Exploration de la Mer, 1971. Fichas de identificación de los estadios larvarios del krill con numerosos dibujos lineales. Archivo PDF, 3 Mb.
Tett, P.: La biología de los eufáusidos , notas de clase de un curso de 2003 sobre Biología Marina de la Universidad Napier .
Tett, P.: Bioluminiscencia , notas de clase de la edición 1999/2000 de ese mismo curso.
Enlaces externos
Wikimedia Commons tiene medios relacionados con Krill.
Wikispecies tiene información relacionada con Euphausia .
Busque krill en Wikcionario, el diccionario libre.
Cámara web del Acuario de Krill de la División Antártica Australiana










{kind=link}