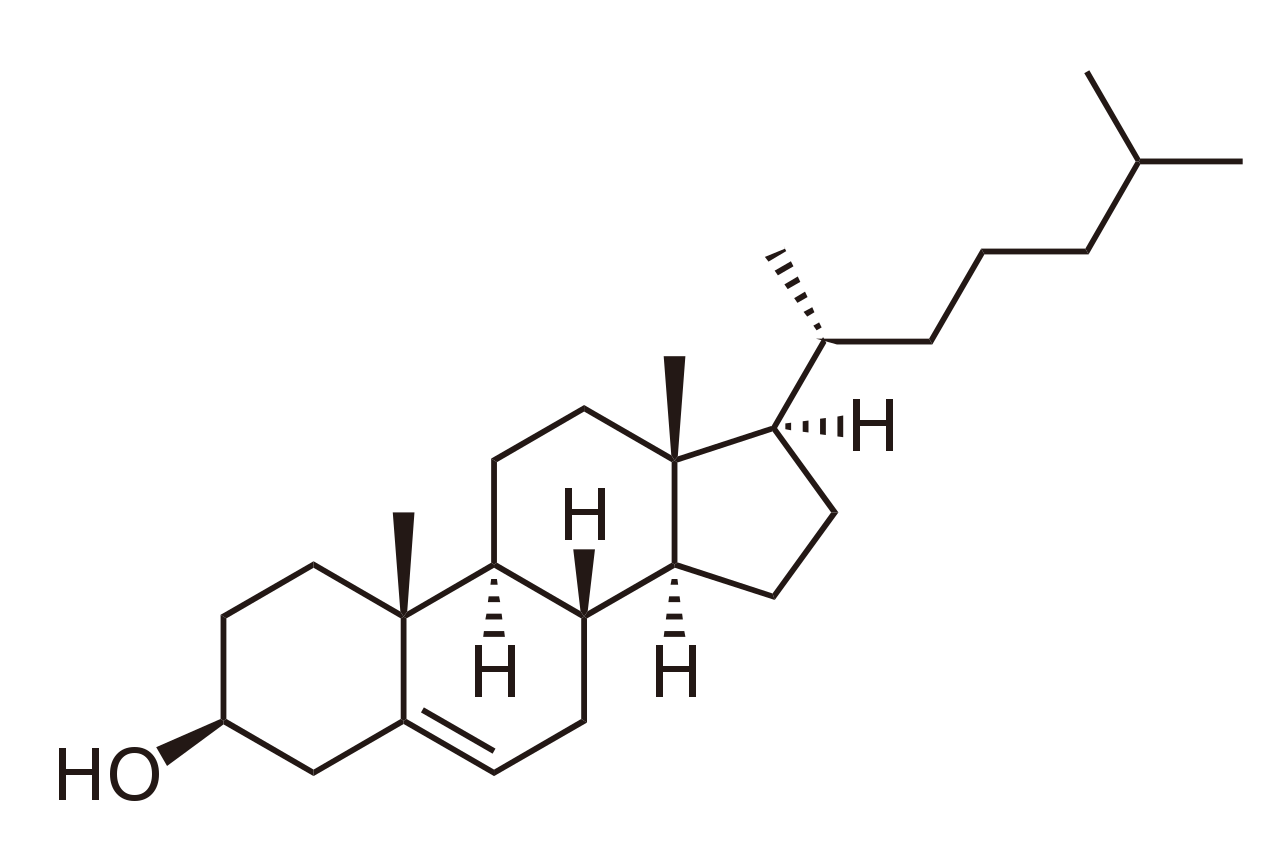
colesterol + 3 NADPH + 3 H + + 3 O 2 ⇄ pregnenolona + 4-metilpentanal + 3 NADP + + 3 H 2 O
P450scc es un miembro de la superfamilia de enzimas del citocromo P450 (familia 11, subfamilia A, polipéptido 1) y está codificado por el gen CYP11A1 . [6]
Nomenclatura
El nombre sistemático de esta clase de enzimas es colesterol, ferredoxina suprarrenal reducida:oxidorreductasa (escisión de cadena lateral) . Otros nombres incluyen:
Localización tisular e intracelular
El nivel más alto del sistema de escisión de la cadena lateral del colesterol se encuentra en la corteza suprarrenal y el cuerpo lúteo . [5] El sistema también se expresa en niveles altos en las células de la teca esteroidogénica en el ovario y en las células de Leydig en el testículo. [5] Durante el embarazo, la placenta también expresa niveles significativos de este sistema enzimático. [7] La P450scc también está presente en niveles mucho más bajos en varios otros tipos de tejidos, incluido el cerebro. [8] En la corteza suprarrenal, la concentración de adrenodoxina es similar a la de P450scc, pero la adrenodoxina reductasa se expresa en niveles más bajos. [9]
Los estudios de inmunofluorescencia utilizando anticuerpos específicos contra las enzimas del sistema P450scc han demostrado que las proteínas se encuentran exclusivamente dentro de las mitocondrias. [10] [11] P450scc se asocia con la membrana mitocondrial interna , mirando hacia el interior (matriz). [12] [13] La adrenodoxina y la adrenodoxina reductasa son proteínas solubles de membrana periférica ubicadas dentro de la matriz mitocondrial que parecen asociarse entre sí principalmente a través de interacciones electrostáticas. [14]
Mecanismo de acción
La P450scc cataliza la conversión de colesterol en pregnenolona en tres reacciones de monooxigenasa. Estas implican dos hidroxilaciones de la cadena lateral del colesterol, que generan, primero, 22R-hidroxicolesterol y luego 20alfa,22R-dihidroxicolesterol. El paso final escinde el enlace entre los carbonos 20 y 22, lo que da como resultado la producción de pregnenolona y aldehído isocaproico.
Cada paso de la monooxigenasa requiere 2 electrones ( equivalentes reductores ). La fuente inicial de los electrones es NADPH. [15] Los electrones se transfieren de NADPH a P450scc a través de dos proteínas de transferencia de electrones: adrenodoxina reductasa [16] y adrenodoxina . [17] [18] Las tres proteínas juntas constituyen el complejo de escisión de la cadena lateral del colesterol.
La participación de tres proteínas en la reacción de escisión de la cadena lateral del colesterol plantea la cuestión de si las tres proteínas funcionan como un complejo ternario como reductasa:adrenodoxina:P450. Tanto los estudios espectroscópicos de la unión de la adrenodoxina a P450scc como los estudios cinéticos en presencia de concentraciones variables de adrenodoxina reductasa demostraron que la reductasa compite con P450scc por la unión a la adrenodoxina. Estos resultados demostraron que la formación de un complejo ternario funcional no es posible. [17] A partir de estos estudios, se concluyó que los sitios de unión de la adrenodoxina a su reductasa y a P450 se superponen y, como consecuencia, la adrenodoxina funciona como una lanzadera de electrones móvil entre la reductasa y P450. [17] Estas conclusiones se han confirmado mediante el análisis estructural del complejo adrenodoxina y P450. [19]
El proceso de transferencia de electrones desde NADPH a P450scc no está estrechamente acoplado; es decir, durante la transferencia de electrones desde la adrenodoxina reductasa a través de la adrenodoxina a P450scc, una cierta porción de los electrones se escapan fuera de la cadena y reaccionan con O 2 , generando radicales superóxido. [20] Las células esteroidogénicas incluyen una variedad diversa de sistemas antioxidantes para hacer frente a los radicales generados por las enzimas esteroidogénicas. [21]
Regulación
En cada célula esteroidogénica, la expresión de las proteínas del sistema P450scc está regulada por el sistema hormonal trófico específico para el tipo celular. [5] En las células de la corteza suprarrenal de la zona fasciculada , la expresión de los ARNm que codifican las tres proteínas P450scc es inducida por la corticotropina (ACTH). [11] [22] Las hormonas tróficas aumentan la expresión del gen CYP11A1 a través de factores de transcripción como el factor esteroidogénico 1 (SF-1), por la isoforma α de la proteína activadora 2 (AP-2) en el ser humano, y muchos otros. [22] [23] La producción de esta enzima es inhibida notablemente por el receptor nuclear DAX-1 . [22]
La p450scc siempre está activa, pero su actividad está limitada por el aporte de colesterol en la membrana interna. Por tanto, el aporte de colesterol a esta membrana (desde la membrana mitocondrial externa ) se considera el verdadero paso limitante de la velocidad de producción de esteroides. Este paso está mediado principalmente por la proteína reguladora aguda esteroidogénica (StAR o STARD1). Tras la estimulación de una célula para que produzca esteroides, la cantidad de StAR disponible para transferir colesterol a la membrana interna limita la velocidad a la que puede ir la reacción (fase aguda). Con una estimulación prolongada (crónica), se cree que el aporte de colesterol ya no es un problema y que la capacidad del sistema para producir esteroides (es decir, el nivel de p450scc en las mitocondrias) es ahora más importante.
La corticotropina (ACTH) es una hormona que se libera desde la hipófisis anterior en respuesta a situaciones de estrés. Un estudio de la capacidad esteroidogénica de la corteza suprarrenal en lactantes con enfermedad respiratoria aguda demostró que efectivamente durante el estado patológico hay un aumento específico de la capacidad esteroidogénica para la síntesis del glucocorticoide cortisol pero no del mineralocorticoide aldosterona o del andrógeno DHEAS que se secretan desde otras zonas de la corteza suprarrenal. [24]
Patología
Las mutaciones en el gen CYP11A1 dan lugar a una deficiencia de hormona esteroide, causando una minoría de casos de la rara y potencialmente mortal enfermedad llamada hiperplasia suprarrenal congénita lipoidea . [25] [26] [27] La deficiencia de CYP11A1 puede provocar hiperpigmentación, hipoglucemia e infecciones recurrentes. [28]
Inhibidores
Los inhibidores de la enzima de escisión de la cadena lateral del colesterol incluyen aminoglutetimida , ketoconazol y mitotano , entre otros. [29] [30] [31]
^ abc ENSG00000288362 GRCh38: Versión 89 de Ensembl: ENSG00000140459, ENSG00000288362 – Ensembl , mayo de 2017
^ abc GRCm38: Lanzamiento de Ensembl 89: ENSMUSG00000032323 – Ensembl , mayo de 2017
^ "Referencia de PubMed humana:". Centro Nacional de Información Biotecnológica, Biblioteca Nacional de Medicina de EE. UU .
^ "Referencia de PubMed sobre ratón". Centro Nacional de Información Biotecnológica, Biblioteca Nacional de Medicina de EE. UU .
^ abcd Hanukoglu I (diciembre de 1992). "Enzimas esteroidogénicas: estructura, función y papel en la regulación de la biosíntesis de hormonas esteroides". Revista de bioquímica y biología molecular de esteroides . 43 (8): 779–804. doi :10.1016/0960-0760(92)90307-5. PMID 22217824. S2CID 112729.
^ "Gen Entrez: CYP11A1 citocromo P450, familia 11, subfamilia A, polipéptido 1".
^ Strauss JF, Martinez F, Kiriakidou M (febrero de 1996). "Síntesis de hormonas esteroides placentarias: características únicas y preguntas sin respuesta". Biology of Reproduction . 54 (2): 303–311. doi : 10.1095/biolreprod54.2.303 . PMID 8788180.
^ Stoffel-Wagner B (diciembre de 2001). "Metabolismo de los neuroesteroides en el cerebro humano". Revista Europea de Endocrinología . 145 (6): 669–679. doi : 10.1530/eje.0.1450669 . PMID 11720889.
^ Hanukoglu I, Hanukoglu Z (mayo de 1986). "Estequiometría de los citocromos mitocondriales P-450, adrenodoxina y adrenodoxina reductasa en la corteza suprarrenal y el cuerpo lúteo. Implicaciones para la organización de la membrana y la regulación génica". Revista Europea de Bioquímica . 157 (1): 27–31. doi : 10.1111/j.1432-1033.1986.tb09633.x . PMID 3011431.
^ Hanukoglu I, Suh BS, Himmelhoch S, Amsterdam A (octubre de 1990). "Inducción y localización mitocondrial de las enzimas del sistema citocromo P450scc en células de la granulosa ovárica normales y transformadas". The Journal of Cell Biology . 111 (4): 1373–1381. doi :10.1083/jcb.111.4.1373. PMC 2116250 . PMID 2170421.
^ ab Hanukoglu I, Feuchtwanger R, Hanukoglu A (noviembre de 1990). "Mecanismo de inducción de enzimas del sistema mitocondrial del citocromo P450 por corticotropina y AMPc en células de la corteza suprarrenal". The Journal of Biological Chemistry . 265 (33): 20602–20608. doi : 10.1016/S0021-9258(17)30545-8 . PMID 2173715.
^ Estudios topológicos de los citocromos P-450scc y P-45011 beta en las membranas mitocondriales internas de la corteza suprarrenal bovina. Efectos de la digestión tríptica controlada. J. Biol. Chem. 1979 254: 10443-8.
^ Farkash Y, Timberg R, Orly J (abril de 1986). "Preparación de antisuero para la escisión de la cadena lateral de colesterol del citocromo P-450 de rata y su uso para la localización ultraestructural de la enzima inmunorreactiva mediante la técnica de proteína A-oro". Endocrinología . 118 (4): 1353–1365. doi : 10.1210/endo-118-4-1353 . PMID 3948785.
^ Hanukoglu I, Privalle CT, Jefcoate CR (mayo de 1981). "Mecanismos de activación iónica de los citocromos mitocondriales suprarrenales P-450scc y P-45011 beta". The Journal of Biological Chemistry . 256 (9): 4329–4335. doi : 10.1016/S0021-9258(19)69437-8 . PMID 6783659.
^ Hanukoglu I, Rapoport R (1995). "Rutas y regulación de la producción de NADPH en mitocondrias esteroidogénicas". Endocrine Research . 21 (1–2): 231–241. doi :10.3109/07435809509030439. PMID 7588385.
^ Hanukoglu I, Gutfinger T, Haniu M, Shively JE (diciembre de 1987). "Aislamiento de un ADNc para la adrenodoxina reductasa (ferredoxina-NADP+ reductasa). Implicaciones para los sistemas mitocondriales del citocromo P-450". Revista Europea de Bioquímica . 169 (3): 449–455. doi : 10.1111/j.1432-1033.1987.tb13632.x . PMID 3691502.
^ abc Hanukoglu I, Jefcoate CR (abril de 1980). "Citocromo mitocondrial P-450scc. Mecanismo de transporte de electrones por adrenodoxina". The Journal of Biological Chemistry . 255 (7): 3057–3061. doi : 10.1016/S0021-9258(19)85851-9 . PMID 6766943.
^ Hanukoglu I, Spitsberg V, Bumpus JA, Dus KM, Jefcoate CR (mayo de 1981). "Citocromo P-450scc mitocondrial suprarrenal. Interacciones entre colesterol y adrenodoxina en equilibrio y durante el recambio". The Journal of Biological Chemistry . 256 (9): 4321–4328. doi : 10.1016/S0021-9258(19)69436-6 . PMID 7217084.
^ Strushkevich N, MacKenzie F, Cherkesova T, Grabovec I, Usanov S, Park HW (junio de 2011). "Base estructural de la biosíntesis de pregnenolona por el sistema monooxigenasa mitocondrial". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 108 (25): 10139–10143. Bibcode :2011PNAS..10810139S. doi : 10.1073/pnas.1019441108 . PMC 3121847 . PMID 21636783.
^ Hanukoglu I, Rapoport R, Weiner L, Sklan D (septiembre de 1993). "Fuga de electrones del sistema mitocondrial NADPH-adrenodoxina reductasa-adrenodoxina-P450scc (escisión de la cadena lateral del colesterol)". Archivos de bioquímica y biofísica . 305 (2): 489–498. doi :10.1006/abbi.1993.1452. PMID 8396893.
^ Hanukoglu I (2006). "Mecanismos de protección antioxidante contra las especies reactivas de oxígeno (ROS) generadas por los sistemas mitocondriales P450 en células esteroidogénicas". Drug Metabolism Reviews . 38 (1–2): 171–196. doi :10.1080/03602530600570040. PMID 16684656. S2CID 10766948.
^ abc Lavoie HA, King SR (agosto de 2009). "Regulación transcripcional de genes esteroidogénicos: STARD1, CYP11A1 y HSD3B". Experimental Biology and Medicine . 234 (8): 880–907. doi :10.3181/0903-MR-97. PMID 19491374. S2CID 5350278.
^ Guo IC, Shih MC, Lan HC, Hsu NC, Hu MC, Chung BC (julio de 2007). "Regulación transcripcional del CYP11A1 humano en gónadas y glándulas suprarrenales". Journal of Biomedical Science . 14 (4): 509–515. doi :10.1007/s11373-007-9177-z. PMID 17594537.
^ Hanukoglu A, Fried D, Nakash I, Hanukoglu I (noviembre de 1995). "Aumentos selectivos en la capacidad esteroidogénica suprarrenal durante la enfermedad respiratoria aguda en lactantes". Revista Europea de Endocrinología . 133 (5): 552–556. doi :10.1530/eje.0.1330552. PMID 7581984. S2CID 44439040.
^ Bhangoo A, Anhalt H, Ten S, King SR (marzo de 2006). "Variaciones fenotípicas en la hiperplasia suprarrenal congénita lipoidea". Pediatric Endocrinology Reviews . 3 (3): 258–271. PMID 16639391.
^ al Kandari H, Katsumata N, Alexander S, Rasoul MA (agosto de 2006). "Mutación homocigótica del gen de la enzima de escisión de la cadena lateral P450 (CYP11A1) en un paciente 46, XY con insuficiencia suprarrenal, inversión sexual completa y agenesia del cuerpo calloso". The Journal of Clinical Endocrinology and Metabolism . 91 (8): 2821–2826. doi :10.1210/jc.2005-2230. PMID 16705068.
^ Kim CJ, Lin L, Huang N, Quigley CA, AvRuskin TW, Achermann JC, Miller WL (marzo de 2008). "Deficiencia suprarrenal y gonadal combinada grave causada por nuevas mutaciones en la enzima de escisión de la cadena lateral del colesterol, P450scc". The Journal of Clinical Endocrinology and Metabolism . 93 (3): 696–702. doi :10.1210/jc.2007-2330. PMC 2266942 . PMID 18182448.
^ FBuonocore F, Achermann JC (2020). "Insuficiencia suprarrenal primaria: nuevas causas genéticas y sus consecuencias a largo plazo". Endocrinología clínica . 92 (1): 11–20. doi :10.1111/cen.14109. PMC 6916405 . PMID 31610036.
^ Becker KL (2001). Principios y práctica de endocrinología y metabolismo. Lippincott Williams & Wilkins. pp. 735–. ISBN978-0-7817-1750-2.
^ Jameson JL, De Groot LJ (18 de mayo de 2010). Endocrinología - Libro electrónico: adultos y niños. Elsevier Health Sciences. págs. 301–302. ISBN978-1-4557-1126-0.
^ Ortiz de Montellano PR (13 de marzo de 2015). Citocromo P450: Estructura, mecanismo y bioquímica. Springer. pp. 851–879. ISBN.978-3-319-12108-6.
Lectura adicional
Helmberg A (agosto de 1993). "Genes gemelos y enfermedades endocrinas: genes CYP21 y CYP11B". Acta Endocrinologica . 129 (2): 97–108. doi :10.1530/acta.0.1290097. PMID 8372604.
Papadopoulos V, Amri H, Boujrad N, Cascio C, Culty M, Garnier M, et al. (enero de 1997). "Receptor periférico de benzodiazepina en el transporte de colesterol y la esteroidogénesis". Esteroides . 62 (1): 21–28. doi : 10.1016/S0039-128X(96)00154-7 . PMID 9029710. S2CID 1977513.
Stocco DM (junio de 2000). "Transferencia intramitocondrial de colesterol". Biochimica et Biophysica Acta (BBA) - Biología molecular y celular de los lípidos . 1486 (1): 184–197. doi :10.1016/S1388-1981(00)00056-1. PMID 10856721.
Kristensen VN, Kure EH, Erikstein B, Harada N, Børresen-Dale A (octubre de 2001). "Susceptibilidad genética y compuestos ambientales similares a los estrógenos". Mutation Research . 482 (1–2): 77–82. doi :10.1016/S0027-5107(01)00212-3. PMID 11535251.
Strauss JF (noviembre de 2003). "Algunas nuevas ideas sobre la fisiopatología y la genética del síndrome de ovario poliquístico". Anales de la Academia de Ciencias de Nueva York . 997 (1): 42–48. Bibcode :2003NYASA.997...42S. doi :10.1196/annals.1290.005. PMID 14644808. S2CID 23559461.
Wada A, Waterman MR (noviembre de 1992). "Identificación mediante mutagénesis dirigida de dos residuos de lisina en el citocromo P450 de escisión de la cadena lateral del colesterol que son esenciales para la unión de la adrenodoxina". The Journal of Biological Chemistry . 267 (32): 22877–22882. doi : 10.1016/S0021-9258(18)50028-4 . PMID 1429635.
Hu MC, Guo IC, Lin JH, Chung BC (marzo de 1991). "Expresión regulada de citocromo P-450scc (enzima de escisión de la cadena lateral del colesterol) en líneas celulares cultivadas detectada por anticuerpos contra la proteína humana expresada en bacterias". The Biochemical Journal . 274 (Pt 3): 813–817. doi :10.1042/bj2740813. PMC 1149983 . PMID 1849407.
Sparkes RS, Klisak I, Miller WL (junio de 1991). "Mapeo regional de genes que codifican enzimas esteroidogénicas humanas: P450scc a 15q23-q24, adrenodoxina a 11q22; adrenodoxina reductasa a 17q24-q25; y P450c17 a 10q24-q25". ADN y biología celular . 10 (5): 359–365. doi :10.1089/dna.1991.10.359. PMID 1863359.
Coghlan VM, Vickery LE (octubre de 1991). "Mutaciones específicas de sitio en la ferredoxina humana que afectan la unión a la ferredoxina reductasa y al citocromo P450scc". The Journal of Biological Chemistry . 266 (28): 18606–18612. doi : 10.1016/S0021-9258(18)55106-1 . PMID 1917982.
Matteson KJ, Chung BC, Urdea MS, Miller WL (abril de 1986). "Estudio de la deficiencia de escisión de la cadena lateral del colesterol (20,22 desmolasa) que causa hiperplasia suprarrenal lipoidea congénita utilizando sondas de oligodesoxirribonucleótidos P450scc de secuencia bovina". Endocrinología . 118 (4): 1296–1305. doi :10.1210/endo-118-4-1296. PMID 2419119.
Chung BC, Matteson KJ, Voutilainen R, Mohandas TK, Miller WL (diciembre de 1986). "Enzima de escisión de la cadena lateral del colesterol humano, P450scc: clonación de ADNc, asignación del gen al cromosoma 15 y expresión en la placenta". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 83 (23): 8962–8966. Bibcode :1986PNAS...83.8962C. doi : 10.1073/pnas.83.23.8962 . PMC 387054 . PMID 3024157.
Morohashi K, Sogawa K, Omura T, Fujii-Kuriyama Y (abril de 1987). "Estructura genética del citocromo P-450 (SCC) humano, colesterol desmolasa". Journal of Biochemistry . 101 (4): 879–887. doi :10.1093/oxfordjournals.jbchem.a121955. PMID 3038854.
Maruyama K, Sugano S (enero de 1994). "Oligo-capping: un método simple para reemplazar la estructura de capuchón de los ARNm eucariotas con oligorribonucleótidos". Gene . 138 (1–2): 171–174. doi :10.1016/0378-1119(94)90802-8. PMID 8125298.
Gharani N, Waterworth DM, Batty S, White D, Gilling-Smith C, Conway GS, et al. (marzo de 1997). "Asociación del gen de síntesis de esteroides CYP11a con el síndrome de ovario poliquístico y el hiperandrogenismo". Human Molecular Genetics . 6 (3): 397–402. doi : 10.1093/hmg/6.3.397 . PMID 9147642.
Suzuki Y, Yoshitomo-Nakagawa K, Maruyama K, Suyama A, Sugano S (octubre de 1997). "Construcción y caracterización de una biblioteca de ADNc enriquecida en longitud completa y enriquecida en el extremo 5'". Gene . 200 (1–2): 149–156. doi :10.1016/S0378-1119(97)00411-3. PMID 9373149.
Hukkanen J, Mäntylä M, Kangas L, Wirta P, Hakkola J, Paakki P, et al. (febrero de 1998). "Expresión de genes del citocromo P450 que codifican enzimas activas en el metabolismo del tamoxifeno en el endometrio uterino humano". Farmacología y Toxicología . 82 (2): 93–97. doi :10.1111/j.1600-0773.1998.tb01404.x. PMID 9498238.
Zhou Z, Shackleton CH, Pahwa S, White PC, Speiser PW (marzo de 1998). "Metabolismo de esteroides sexuales prominentes en linfocitos humanos". Endocrinología molecular y celular . 138 (1–2): 61–69. doi : 10.1016/S0303-7207(98)00052-5 . PMID 9685215. S2CID 20490901.
Síntesis de hormonas esteroides
Esteroidogénesis , que muestra la enzima de escisión de la cadena lateral del colesterol en la parte superior.Síntesis de hormonas esteroides




{kind=link}