_with_its_prey.jpg/1280px-Polar_bear_(Ursus_maritimus)_with_its_prey.jpg)

La depredación es una interacción biológica en la que un organismo, el depredador , mata y se come a otro organismo, su presa . Es parte de una familia de comportamientos alimentarios comunes que incluyen el parasitismo y la microdepredación (que generalmente no matan al huésped ) y el parasitoidismo (que siempre lo hace, eventualmente). Es diferente de la carroña de presas muertas, aunque muchos depredadores también carroñean ; se superpone con la herbivoría , ya que los depredadores de semillas y los frugívoros destructivos son depredadores.
Los depredadores pueden buscar o perseguir activamente a sus presas o esperarlas, a menudo escondidos. Cuando detectan una presa, el depredador evalúa si atacarla. Esto puede implicar una emboscada o una depredación por persecución , a veces después de acechar a la presa. Si el ataque tiene éxito, el depredador mata a la presa, le quita las partes no comestibles, como el caparazón o las espinas, y se la come.
Los depredadores están adaptados y a menudo altamente especializados para la caza, con sentidos agudos como la vista , el oído o el olfato . Muchos animales depredadores , tanto vertebrados como invertebrados , tienen garras o mandíbulas afiladas para agarrar, matar y cortar a sus presas. Otras adaptaciones incluyen el sigilo y el mimetismo agresivo que mejoran la eficiencia de la caza.
La depredación tiene un poderoso efecto selectivo sobre las presas, y estas desarrollan adaptaciones antidepredadoras, como coloración de advertencia , llamadas de alarma y otras señales , camuflaje , imitación de especies bien defendidas y espinas y sustancias químicas defensivas. A veces, depredadores y presas se encuentran en una carrera armamentista evolutiva , un ciclo de adaptaciones y contraadaptaciones. La depredación ha sido un importante motor de la evolución al menos desde el período Cámbrico .

En el nivel más básico, los depredadores matan y comen a otros organismos. Sin embargo, el concepto de depredación es amplio, se define de manera diferente en diferentes contextos e incluye una amplia variedad de métodos de alimentación; además, algunas relaciones que resultan en la muerte de la presa no necesariamente se denominan depredación. Un parasitoide , como una avispa icneumón , pone sus huevos en o sobre su anfitrión; los huevos eclosionan en larvas, que se comen al anfitrión, y este inevitablemente muere. Los zoólogos generalmente llaman a esto una forma de parasitismo , aunque convencionalmente se piensa que los parásitos no matan a sus anfitriones. Un depredador puede definirse como diferente de un parasitoide en que tiene muchas presas, capturadas a lo largo de su vida, mientras que la larva de un parasitoide tiene solo una, o al menos tiene su suministro de alimento provisto para ella en una sola ocasión. [1] [2]

Existen otros casos difíciles y limítrofes. Los microdepredadores son animales pequeños que, como los depredadores, se alimentan completamente de otros organismos; incluyen pulgas y mosquitos que consumen sangre de animales vivos, y pulgones que consumen savia de plantas vivas. Sin embargo, como normalmente no matan a sus huéspedes, ahora se los suele considerar parásitos. [3] [4] Los animales que pastan en fitoplancton o esteras de microbios son depredadores, ya que consumen y matan a sus organismos alimentarios, mientras que los herbívoros que ramonean hojas no lo son, ya que sus plantas alimenticias generalmente sobreviven al ataque. [5] Cuando los animales comen semillas ( depredación de semillas o granivoría ) o huevos ( depredación de huevos ), están consumiendo organismos vivos completos, lo que por definición los convierte en depredadores. [6] [7] [8]
Los carroñeros , organismos que solo se alimentan de organismos que se encuentran ya muertos, no son depredadores, pero muchos depredadores como el chacal y la hiena carroñean cuando surge la oportunidad. [9] [10] [5] Entre los invertebrados, las avispas sociales como las avispas amarillas son a la vez cazadoras y carroñeras de otros insectos. [11]
Si bien los ejemplos de depredadores entre mamíferos y aves son bien conocidos, [12] se pueden encontrar depredadores en una amplia gama de taxones, incluidos los artrópodos. Son comunes entre los insectos, incluidas las mantis, las libélulas , las crisopas y las moscas escorpión . En algunas especies, como la mosca aliso , solo las larvas son depredadoras (los adultos no comen). Las arañas son depredadoras, así como otros invertebrados terrestres como los escorpiones ; ciempiés ; algunos ácaros , caracoles y babosas ; nematodos ; y gusanos planarios . [13] En entornos marinos, la mayoría de los cnidarios (p. ej., medusas , hidroides ), ctenóforos (medusas peine), equinodermos (p. ej., estrellas de mar , erizos de mar , dólares de arena y pepinos de mar ) y gusanos planos son depredadores. [14] Entre los crustáceos , las langostas , los cangrejos , los camarones y los percebes son depredadores, [15] y a su vez los crustáceos son presa de casi todos los cefalópodos (incluidos pulpos , calamares y sepias ). [16]

La depredación de semillas está restringida a mamíferos, aves e insectos, pero se encuentra en casi todos los ecosistemas terrestres. [8] [6] La depredación de huevos incluye tanto a depredadores de huevos especializados, como algunas serpientes colúbridas , como a depredadores generalistas, como zorros y tejones, que toman huevos de manera oportunista cuando los encuentran. [17] [18] [19]
Algunas plantas, como la planta carnívora , la venus atrapamoscas y la drosera , son carnívoras y consumen insectos . [12] Los métodos de depredación de las plantas varían mucho, pero a menudo implican una trampa de comida, estimulación mecánica e impulsos eléctricos para finalmente atrapar y consumir a su presa. [20] Algunos hongos carnívoros capturan nematodos utilizando trampas activas en forma de anillos constrictores o trampas pasivas con estructuras adhesivas. [21]
Muchas especies de protozoos ( eucariotas ) y bacterias ( procariotas ) se alimentan de otros microorganismos; el modo de alimentación es evidentemente antiguo y evolucionó muchas veces en ambos grupos. [22] [12] [23] Entre el zooplancton de agua dulce y marino , ya sea unicelular o multicelular, el pastoreo depredador de fitoplancton y zooplancton más pequeño es común, y se encuentra en muchas especies de nanoflagelados , dinoflagelados , ciliados , rotíferos , una amplia gama de larvas de animales de meroplancton y dos grupos de crustáceos, a saber, copépodos y cladóceros . [24]

Para alimentarse, un depredador debe buscar, perseguir y matar a su presa. Estas acciones forman un ciclo de búsqueda de alimento . [26] [27] El depredador debe decidir dónde buscar a la presa en función de su distribución geográfica; y una vez que ha localizado a la presa, debe evaluar si la persigue o espera una mejor opción. Si elige la persecución, sus capacidades físicas determinan el modo de persecución (por ejemplo, emboscada o persecución). [28] [29] Una vez capturada la presa, es posible que también deba gastar energía en manipularla (por ejemplo, matarla, quitarle el caparazón o las espinas e ingerirla). [25] [26]
Los depredadores pueden elegir entre distintos modos de búsqueda, desde sentarse y esperar hasta la búsqueda activa o amplia . [30] [25] [31] [32] El método de sentarse y esperar es el más adecuado si las presas son densas y móviles, y el depredador tiene bajos requerimientos de energía. [30] La búsqueda amplia gasta más energía y se utiliza cuando la presa es sedentaria o está escasamente distribuida. [28] [30] Existe un continuo de modos de búsqueda con intervalos entre períodos de movimiento que van desde segundos hasta meses. Los tiburones, peces luna , aves insectívoras y musarañas casi siempre se mueven, mientras que las arañas tejedoras de telarañas, los invertebrados acuáticos, las mantis religiosas y los cernícalos rara vez se mueven. Entre tanto, los chorlitos y otras aves playeras , los peces de agua dulce, incluidas las percas , y las larvas de los escarabajos coccinélidos (mariquitas) , alternan entre la búsqueda activa y el escaneo del entorno. [30]

Las distribuciones de presas a menudo están agrupadas, y los depredadores responden buscando parches donde la presa es densa y luego buscando dentro de los parches. [25] Cuando se encuentra alimento en parches, como bancos raros de peces en un océano casi vacío, la etapa de búsqueda requiere que el depredador viaje durante un tiempo sustancial y gaste una cantidad significativa de energía para localizar cada parche de alimento. [33] Por ejemplo, el albatros de ceja negra realiza vuelos de alimentación regularmente a un rango de alrededor de 700 kilómetros (430 millas), hasta un rango de alimentación máximo de 3000 kilómetros (1860 millas) para aves reproductoras que recolectan alimento para sus crías. [a] [34] Con presas estáticas, algunos depredadores pueden aprender ubicaciones de parches adecuados y regresar a ellos a intervalos para alimentarse. [33] La estrategia de alimentación óptima para la búsqueda se ha modelado utilizando el teorema del valor marginal . [35]
Los patrones de búsqueda suelen parecer aleatorios. Uno de ellos es el paseo de Lévy , que suele incluir grupos de pasos cortos con pasos largos ocasionales. Se ajusta bien al comportamiento de una amplia variedad de organismos, incluidas bacterias, abejas, tiburones y cazadores-recolectores humanos. [36] [37]

Una vez que encuentra una presa, el depredador debe decidir si la persigue o sigue buscando. La decisión depende de los costos y beneficios involucrados. Un ave que busca insectos pasa mucho tiempo buscándolos, pero capturarlos y comerlos es rápido y fácil, por lo que la estrategia eficiente para el ave es comer todos los insectos apetitosos que encuentra. Por el contrario, un depredador como un león o un halcón encuentra su presa fácilmente, pero capturarla requiere mucho esfuerzo. En ese caso, el depredador es más selectivo. [28]
Uno de los factores a tener en cuenta es el tamaño. Una presa demasiado pequeña puede no merecer la pena por la cantidad de energía que proporciona. Si es demasiado grande, puede resultar demasiado difícil capturarla. Por ejemplo, una mantis captura a sus presas con sus patas delanteras, que están optimizadas para atrapar presas de un determinado tamaño. Las mantis son reacias a atacar a presas que estén lejos de ese tamaño. Existe una correlación positiva entre el tamaño de un depredador y su presa. [28]
Un depredador puede evaluar una zona y decidir si pasa tiempo buscando presas en ella. [25] Esto puede implicar cierto conocimiento de las preferencias de la presa; por ejemplo, las mariquitas pueden elegir una zona de vegetación adecuada para sus presas, los pulgones . [38]
Para capturar presas, los depredadores tienen un espectro de modos de persecución que van desde la persecución abierta ( depredación por persecución ) hasta un ataque repentino a una presa cercana ( depredación por emboscada ). [25] [39] [12] Otra estrategia entre la emboscada y la persecución es la interceptación balística , donde un depredador observa y predice el movimiento de una presa y luego lanza su ataque en consecuencia. [40]
Los depredadores de emboscada o de acecho son animales carnívoros que capturan presas por sigilo o sorpresa. En los animales, la depredación por emboscada se caracteriza por el depredador explorando el entorno desde una posición oculta hasta que detecta una presa y luego ejecutando rápidamente un ataque sorpresa fijo. [41] [40] Los depredadores vertebrados de emboscada incluyen ranas, peces como el tiburón ángel , el lucio del norte y el pez sapo oriental . [40] [42] [43] [44] Entre los muchos depredadores invertebrados de emboscada se encuentran las arañas de trampilla y las arañas cangrejo australianas en la tierra y los camarones mantis en el mar. [41] [45] [46] Los depredadores de emboscada a menudo construyen una madriguera en la que esconderse, mejorando el ocultamiento a costa de reducir su campo de visión. Algunos depredadores de emboscada también usan señuelos para atraer presas dentro del alcance de ataque. [40] El movimiento de captura debe ser rápido para atrapar a la presa, dado que el ataque no es modificable una vez lanzado. [40]

La interceptación balística es una estrategia en la que un depredador observa el movimiento de una presa, predice su movimiento, calcula una ruta de interceptación y luego ataca a la presa en esa ruta. Esto difiere de la depredación por emboscada en que el depredador ajusta su ataque según cómo se mueve la presa. [40] La interceptación balística implica un breve período de planificación, lo que le da a la presa la oportunidad de escapar. Algunas ranas esperan hasta que las serpientes hayan comenzado su ataque antes de saltar, lo que reduce el tiempo disponible para que la serpiente recalibre su ataque y maximiza el ajuste angular que la serpiente necesitaría hacer para interceptar a la rana en tiempo real. [40] Los depredadores balísticos incluyen insectos como las libélulas y vertebrados como el pez arquero (que ataca con un chorro de agua), los camaleones (que atacan con la lengua) y algunas serpientes colúbridas . [40]
En la depredación por persecución, los depredadores persiguen a una presa que huye. Si la presa huye en línea recta, la captura depende únicamente de que el depredador sea más rápido que la presa. [40] Si la presa maniobra girando mientras huye, el depredador debe reaccionar en tiempo real para calcular y seguir una nueva ruta de intercepción, como por ejemplo mediante navegación paralela , a medida que se acerca a la presa. [40] Muchos depredadores de persecución utilizan el camuflaje para acercarse a la presa lo más cerca posible sin ser observados ( acecho ) antes de comenzar la persecución. [40] Los depredadores de persecución incluyen mamíferos terrestres como humanos, perros salvajes africanos, hienas manchadas y lobos; depredadores marinos como delfines, orcas y muchos peces depredadores, como el atún; [47] [48] aves depredadoras (rapaz) como halcones; e insectos como libélulas . [49]
Una forma extrema de persecución es la caza de resistencia o persistencia , en la que el depredador cansa a la presa siguiéndola a lo largo de una gran distancia, a veces durante horas. El método es utilizado por cazadores-recolectores humanos y por cánidos como los perros salvajes africanos y los perros domésticos. El perro salvaje africano es un depredador de persistencia extrema, que cansa a las presas individuales siguiéndolas durante muchos kilómetros a una velocidad relativamente baja. [50]
Una forma especializada de depredación por persecución es la alimentación por embestida de las ballenas barbadas . Estos depredadores marinos de gran tamaño se alimentan de plancton , especialmente krill , sumergiéndose y nadando activamente en concentraciones de plancton, para luego tomar una gran cantidad de agua y filtrarla a través de sus placas barbadas . [ 51] [52]
Los depredadores de persecución pueden ser sociales , como el león y el lobo que cazan en grupos, o solitarios. [2]
Una vez que el depredador ha capturado a la presa, debe manipularla: con mucho cuidado si la presa es peligrosa para comer, como si posee espinas afiladas o venenosas, como en muchos peces presa. Algunos bagres , como los Ictaluridae, tienen espinas en la espalda (dorsal) y el vientre (pectoral) que se bloquean en la posición erecta; cuando el bagre se agita cuando es capturado, estas podrían perforar la boca del depredador, posiblemente fatalmente. Algunas aves que se alimentan de peces, como el águila pescadora, evitan el peligro de las espinas desgarrando a su presa antes de comérsela. [53]
En la depredación social, un grupo de depredadores coopera para matar a sus presas. Esto permite matar criaturas más grandes que las que podrían dominar individualmente; por ejemplo, las hienas y los lobos colaboran para atrapar y matar herbívoros tan grandes como los búfalos, y los leones incluso cazan elefantes. [54] [55] [56] También puede hacer que las presas estén más fácilmente disponibles a través de estrategias como hacerlas salir volando y arrearlas a un área más pequeña. Por ejemplo, cuando bandadas mixtas de aves buscan comida, las aves que van al frente hacen salir a los insectos que son atrapados por las aves que están detrás. Los delfines giradores forman un círculo alrededor de un banco de peces y se mueven hacia adentro, concentrando a los peces en un factor de 200. [57] Al cazar socialmente, los chimpancés pueden atrapar monos colobos que escaparían fácilmente de un cazador individual, mientras que los halcones de Harris cooperantes pueden atrapar conejos. [54] [58]

Los depredadores de diferentes especies a veces cooperan para atrapar presas. En los arrecifes de coral , cuando peces como el mero y la trucha coral detectan presas que son inaccesibles para ellos, envían señales a morenas gigantes , lábridos Napoleón o pulpos . Estos depredadores pueden acceder a pequeñas grietas y hacer salir a la presa. [59] [60] Se sabe que las orcas ayudan a los balleneros a cazar ballenas barbadas . [61]
La caza social permite a los depredadores enfrentarse a una gama más amplia de presas, pero con el riesgo de competir por el alimento capturado. Los depredadores solitarios tienen más posibilidades de comer lo que capturan, al precio de un mayor gasto de energía para capturarlo y un mayor riesgo de que la presa escape. [62] [63] Los depredadores de emboscada suelen ser solitarios para reducir el riesgo de convertirse ellos mismos en presas. [64] De los 245 miembros terrestres de los carnívoros (el grupo que incluye a los gatos, perros y osos), 177 son solitarios; y 35 de los 37 gatos salvajes son solitarios, [65] incluyendo al puma y al guepardo. [62] [2] Sin embargo, el puma solitario permite que otros pumas compartan una presa, [66] y el coyote puede ser solitario o social. [67] Otros depredadores solitarios incluyen al lucio del norte, [68] las arañas lobo y todas las miles de especies de avispas solitarias entre los artrópodos, [69] [70] y muchos microorganismos y zooplancton . [22] [71]
Bajo la presión de la selección natural , los depredadores han desarrollado una variedad de adaptaciones físicas para detectar, atrapar, matar y digerir a sus presas, entre ellas velocidad, agilidad, sigilo, sentidos agudos, garras, dientes, filtros y sistemas digestivos adecuados. [72]
Para detectar presas , los depredadores tienen una visión , un olfato o un oído bien desarrollados . [12] Depredadores tan diversos como los búhos y las arañas saltadoras tienen ojos orientados hacia adelante, lo que proporciona una visión binocular precisa en un campo de visión relativamente estrecho, mientras que los animales presa a menudo tienen una visión panorámica menos aguda. Animales como los zorros pueden oler a sus presas incluso cuando están ocultas bajo 2 pies (60 cm) de nieve o tierra. Muchos depredadores tienen un oído agudo y algunos, como los murciélagos ecolocalizadores, cazan exclusivamente mediante el uso activo o pasivo del sonido. [73]
Los depredadores, incluidos los grandes felinos , las aves rapaces y las hormigas, comparten mandíbulas poderosas, dientes afilados o garras que utilizan para atrapar y matar a sus presas. Algunos depredadores, como las serpientes y las aves que se alimentan de peces, como las garzas y los cormoranes, se tragan a sus presas enteras; algunas serpientes pueden desencajar sus mandíbulas para poder tragar presas grandes, mientras que las aves que se alimentan de peces tienen picos largos en forma de lanza que utilizan para apuñalar y agarrar presas que se mueven rápidamente y son resbaladizas. [73] Los peces y otros depredadores han desarrollado la capacidad de aplastar o abrir las conchas blindadas de los moluscos. [74]
Muchos depredadores tienen una constitución fuerte y pueden atrapar y matar animales más grandes que ellos; esto se aplica tanto a pequeños depredadores como hormigas y musarañas como a carnívoros grandes y visiblemente musculosos como el puma y el león . [73] [2] [75]
.JPG/1280px-Ursus_arctos_01_MWNH_145_(cropped).JPG)

Los depredadores suelen estar altamente especializados en su dieta y comportamiento de caza; por ejemplo, el lince euroasiático solo caza pequeños ungulados . [76] Otros, como los leopardos , son generalistas más oportunistas y se alimentan de al menos 100 especies. [77] [78] Los especialistas pueden estar muy adaptados a capturar su presa preferida, mientras que los generalistas pueden ser más capaces de cambiar a otra presa cuando un objetivo preferido es escaso. Cuando las presas tienen una distribución agrupada (desigual), se predice que la estrategia óptima para el depredador será más especializada, ya que las presas son más visibles y se pueden encontrar más rápidamente; [79] esto parece ser correcto para los depredadores de presas inmóviles, pero es dudoso con presas móviles. [80]
En la depredación selectiva por tamaño, los depredadores seleccionan presas de un tamaño determinado. [81] Las presas grandes pueden resultar problemáticas para un depredador, mientras que las presas pequeñas pueden resultar difíciles de encontrar y, en cualquier caso, proporcionar una recompensa menor. Esto ha llevado a una correlación entre el tamaño de los depredadores y sus presas. El tamaño también puede actuar como refugio para las presas grandes. Por ejemplo, los elefantes adultos están relativamente a salvo de la depredación de los leones, pero los jóvenes son vulnerables. [82]
Los miembros de la familia de los felinos , como el leopardo de las nieves (tierras altas sin árboles), el tigre (llanuras cubiertas de hierba, pantanos de juncos), el ocelote (bosque), el gato pescador (matorrales ribereños) y el león (llanuras abiertas) se camuflan con coloraciones y patrones disruptivos que se adaptan a sus hábitats. [83]
En el mimetismo agresivo , ciertos depredadores, incluidos insectos y peces, hacen uso de la coloración y el comportamiento para atraer presas. Las luciérnagas Photuris hembras , por ejemplo, copian las señales luminosas de otras especies, atrayendo así a las luciérnagas macho, que capturan y comen. [84] Las mantis de las flores son depredadores de emboscada; camufladas como flores, como las orquídeas , atraen a sus presas y las capturan cuando están lo suficientemente cerca. [85] Los peces sapo están extremadamente bien camuflados y atraen activamente a sus presas para que se acerquen usando una esca , un cebo en el extremo de un apéndice en forma de varilla en la cabeza, que agitan suavemente para imitar a un animal pequeño, engullendo a la presa en un movimiento extremadamente rápido cuando está dentro del alcance. [86]
Muchos depredadores más pequeños, como las medusas, utilizan veneno para someter a sus presas [87] y el veneno también puede ayudar a la digestión (como es el caso de las serpientes de cascabel y algunas arañas ). [88] [89] La serpiente marina jaspeada que se ha adaptado a la depredación de huevos tiene glándulas venenosas atrofiadas y el gen de su toxina de tres dedos contiene una mutación (la eliminación de dos nucleótidos ) que la inactiva. Estos cambios se explican por el hecho de que su presa no necesita ser sometida. [90]

Varios grupos de peces depredadores tienen la capacidad de detectar, rastrear y, a veces, como en el caso del rayo eléctrico , incapacitar a sus presas al detectar y generar campos eléctricos . [91] [92] [93] El órgano eléctrico se deriva de tejido nervioso o muscular modificado. [94]
Las adaptaciones fisiológicas a la depredación incluyen la capacidad de las bacterias depredadoras de digerir el complejo polímero de peptidoglicano de las paredes celulares de las bacterias que cazan. [23] Los vertebrados carnívoros de las cinco clases principales (peces, anfibios, reptiles, aves y mamíferos) tienen tasas relativas más bajas de transporte de azúcar a aminoácidos que los herbívoros u omnívoros, presumiblemente porque adquieren una gran cantidad de aminoácidos de las proteínas animales en su dieta. [95]
Para contrarrestar la depredación, las presas han desarrollado defensas para usar en cada etapa de un ataque. [96] [12] Pueden tratar de evitar ser detectadas, [97] como mediante el uso del camuflaje y el mimetismo . [98] Pueden detectar depredadores [99] y advertir a otros de su presencia. [100] [101] Si son detectadas, pueden tratar de evitar ser el objetivo de un ataque, por ejemplo, señalando que son tóxicas o desagradables , [102] [103] [104] señalando que una persecución no sería rentable, [105] [106] o formando grupos. [107] [108] Si se convierten en un objetivo, pueden tratar de defenderse del ataque con defensas como armaduras, púas , desagradables o acoso; [109] [110] [111] y a menudo pueden escapar de un ataque en curso asustando al depredador, [112] [113] [114] haciéndose los muertos , desprendiéndose de partes del cuerpo como la cola, o simplemente huyendo. [115] [116]

Los depredadores y sus presas son enemigos naturales, y muchas de sus adaptaciones parecen diseñadas para contrarrestarse entre sí. Por ejemplo, los murciélagos tienen sofisticados sistemas de ecolocalización para detectar insectos y otras presas, y los insectos han desarrollado una variedad de defensas, incluida la capacidad de escuchar los llamados de ecolocalización. [117] [118] Muchos depredadores de persecución que corren sobre la tierra, como los lobos, han desarrollado extremidades largas en respuesta a la mayor velocidad de sus presas. [119] Sus adaptaciones se han caracterizado como una carrera armamentista evolutiva , un ejemplo de la coevolución de dos especies. [120] En una visión de la evolución centrada en los genes , se puede pensar que los genes del depredador y la presa compiten por el cuerpo de la presa. [120] Sin embargo, el principio de "vida-cena" de Dawkins y Krebs predice que esta carrera armamentista es asimétrica: si un depredador no logra atrapar a su presa, pierde su cena, mientras que si tiene éxito, la presa pierde su vida. [120]

La metáfora de una carrera armamentística implica avances cada vez mayores en ataque y defensa. Sin embargo, estas adaptaciones tienen un costo; por ejemplo, las patas más largas tienen un mayor riesgo de romperse, [121] mientras que la lengua especializada del camaleón, con su capacidad de actuar como un proyectil, es inútil para lamer el agua, por lo que el camaleón debe beber el rocío de la vegetación. [122]
El principio de la "vida-cena" ha sido criticado por múltiples motivos. El grado de asimetría en la selección natural depende en parte de la heredabilidad de los rasgos adaptativos. [122] Además, si un depredador pierde suficientes cenas, también perderá su vida. [121] [122] Por otro lado, el costo de adaptación de una determinada cena perdida es impredecible, ya que el depredador puede encontrar rápidamente una presa mejor. Además, la mayoría de los depredadores son generalistas, lo que reduce el impacto de una determinada adaptación de una presa en un depredador. Dado que la especialización es causada por la coevolución depredador-presa, la rareza de los especialistas puede implicar que las carreras armamentistas depredador-presa son raras. [122]
Es difícil determinar si las adaptaciones dadas son realmente el resultado de la coevolución, donde una adaptación de la presa da lugar a una adaptación del depredador que es contrarrestada por una adaptación adicional en la presa. Una explicación alternativa es la escalada , donde los depredadores se adaptan a los competidores, sus propios depredadores o presas peligrosas. [123] Las adaptaciones aparentes a la depredación también pueden haber surgido por otras razones y luego haber sido cooptadas para el ataque o la defensa. En algunos de los insectos presa de los murciélagos, la audición evolucionó antes de que aparecieran los murciélagos y se utilizó para escuchar señales utilizadas para la defensa territorial y el apareamiento. [124] Su audición evolucionó en respuesta a la depredación de los murciélagos, pero el único ejemplo claro de adaptación recíproca en los murciélagos es la ecolocalización sigilosa. [125]
Una carrera armamentista más simétrica puede ocurrir cuando las presas son peligrosas, tienen espinas, púas, toxinas o veneno que pueden dañar al depredador. El depredador puede responder con evitación, lo que a su vez impulsa la evolución del mimetismo. La evitación no es necesariamente una respuesta evolutiva, ya que generalmente se aprende de malas experiencias con la presa. Sin embargo, cuando la presa es capaz de matar al depredador (como puede hacerlo una serpiente coral con su veneno), no hay oportunidad de aprendizaje y la evitación debe ser hereditaria. Los depredadores también pueden responder a presas peligrosas con contraadaptaciones. En el oeste de América del Norte, la serpiente de liga común ha desarrollado una resistencia a la toxina en la piel del tritón de piel áspera . [122]
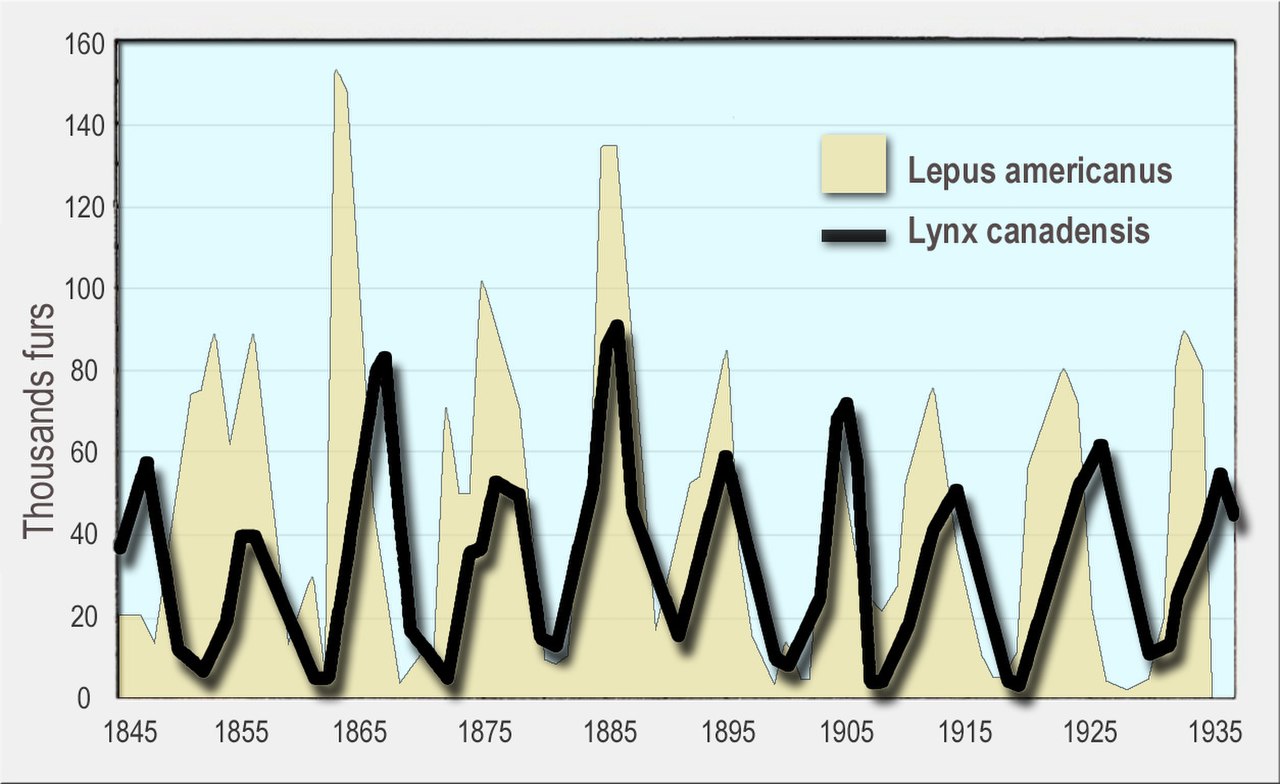
Los depredadores afectan a sus ecosistemas no sólo directamente al comer sus propias presas, sino también por medios indirectos, como la reducción de la depredación por parte de otras especies o la alteración del comportamiento de búsqueda de alimento de un herbívoro, como en el caso del efecto de los lobos sobre la vegetación de ribera o de las nutrias marinas sobre los bosques de algas marinas en la biodiversidad. Esto puede explicar los efectos de la dinámica de poblaciones, como los ciclos observados en el lince y la liebre de raquetas de nieve. [126] [127] [128]
Una forma de clasificar a los depredadores es por nivel trófico . Los carnívoros que se alimentan de herbívoros son consumidores secundarios; sus depredadores son consumidores terciarios, y así sucesivamente. [129] En la cima de esta cadena alimentaria se encuentran los depredadores de ápice como los leones . [130] Sin embargo, muchos depredadores comen de múltiples niveles de la cadena alimentaria; un carnívoro puede comer tanto consumidores secundarios como terciarios. [131] Esto significa que muchos depredadores deben lidiar con la depredación intragremial , donde otros depredadores los matan y se los comen. Por ejemplo, los coyotes compiten con zorros grises y gatos monteses y, a veces, los matan . [132]
La transferencia trófica dentro de un ecosistema se refiere al transporte de energía y nutrientes como resultado de la depredación. La energía pasa de un nivel trófico al siguiente a medida que los depredadores consumen materia orgánica del cuerpo de otro organismo . Dentro de cada transferencia, si bien hay usos de energía, también hay pérdidas de energía.
Los niveles tróficos marinos varían según la localidad y el tamaño de los productores primarios . Generalmente hay hasta seis niveles tróficos en el océano abierto, cuatro sobre las plataformas continentales y alrededor de tres en las zonas de surgencia. [133] Por ejemplo, un hábitat marino con cinco niveles tróficos podría representarse de la siguiente manera: Herbívoros (se alimentan principalmente de fitoplancton ); Carnívoros (se alimentan principalmente de otro zooplancton /animales); Detritívoros (se alimentan principalmente de materia orgánica muerta/ detritos ); Omnívoros (se alimentan de una dieta mixta de fito- y zooplancton y detritos); y Mixótrofos que combinan la autotrofia (usando la energía de la luz para crecer sin ingerir ningún compuesto orgánico o nutriente adicional) con la heterotrofia (alimentándose de otras plantas y animales para obtener energía y nutrientes: herbívoros, omnívoros y carnívoros, y detritívoros).
La eficiencia de transferencia trófica mide la eficacia con la que la energía se transfiere o pasa a través de los niveles tróficos superiores de la red alimentaria marina . A medida que la energía asciende por los niveles tróficos, disminuye debido al calor, los desechos y los procesos metabólicos naturales que ocurren cuando los depredadores consumen a sus presas. El resultado es que solo alrededor del 10% de la energía en cualquier nivel trófico se transfiere al siguiente nivel. Esto se conoce a menudo como "la regla del 10%", que limita el número de niveles tróficos que un ecosistema individual es capaz de soportar. [134]
Los depredadores pueden aumentar la biodiversidad de las comunidades al impedir que una sola especie se vuelva dominante. Estos depredadores se conocen como especies clave y pueden tener una profunda influencia en el equilibrio de los organismos en un ecosistema particular . [135] La introducción o eliminación de este depredador, o los cambios en su densidad de población, pueden tener efectos en cascada drásticos en el equilibrio de muchas otras poblaciones en el ecosistema. Por ejemplo, los animales que pastan en un pastizal pueden impedir que una sola especie dominante tome el control. [136]
_horiz.jpg/1280px-Fig._1_-Riparian_willow_recovery_(26485120926)_horiz.jpg)
La eliminación de los lobos del Parque Nacional de Yellowstone tuvo un profundo impacto en la pirámide trófica . En esa zona, los lobos son especies clave y depredadores máximos. Sin depredación, los herbívoros comenzaron a sobrepastorear muchas especies de ramoneos leñosos, afectando a las poblaciones de plantas de la zona. Además, los lobos a menudo impedían que los animales pastaran cerca de los arroyos, protegiendo así las fuentes de alimento de los castores . La eliminación de los lobos tuvo un efecto directo en la población de castores, ya que su hábitat se convirtió en territorio para el pastoreo. El aumento del ramoneo de sauces y coníferas a lo largo del arroyo Blacktail debido a la falta de depredación provocó una incisión en el canal porque la población reducida de castores ya no podía frenar el agua y mantener el suelo en su lugar. De este modo, se demostró que los depredadores eran de vital importancia en el ecosistema. [137]
En ausencia de depredadores, la población de una especie puede crecer exponencialmente hasta aproximarse a la capacidad de sustentación del ambiente. [138] Los depredadores limitan el crecimiento de las presas tanto consumiéndolas como modificando su comportamiento. [139] Los aumentos o disminuciones en la población de presas también pueden conducir a aumentos o disminuciones en el número de depredadores, por ejemplo, a través de un aumento en el número de crías que tienen.
Se han observado fluctuaciones cíclicas en las poblaciones de depredadores y presas, a menudo con desfases entre los ciclos de depredador y presa. Un ejemplo bien conocido es el de la liebre de raquetas de nieve y el lince . En una amplia extensión de bosques boreales de Alaska y Canadá, las poblaciones de liebres fluctúan casi en sincronía con un período de 10 años, y las poblaciones de linces fluctúan en respuesta. Esto se vio por primera vez en los registros históricos de animales capturados por cazadores de pieles para la Compañía de la Bahía de Hudson durante más de un siglo. [140] [128] [141] [142]
.png/1280px-Lotka-Volterra_model_(1.1,_0.4,_0.4,_0.1).png)
Un modelo simple de un sistema con una especie de depredador y una de presa, las ecuaciones de Lotka-Volterra , predicen ciclos poblacionales. [143] Sin embargo, los intentos de reproducir las predicciones de este modelo en el laboratorio a menudo han fracasado; por ejemplo, cuando el protozoo Didinium nasutum se agrega a un cultivo que contiene su presa, Paramecium caudatum , esta última a menudo se ve llevada a la extinción. [144]
Las ecuaciones de Lotka-Volterra se basan en varios supuestos simplificadores y son estructuralmente inestables , lo que significa que cualquier cambio en las ecuaciones puede estabilizar o desestabilizar la dinámica. [145] [146] Por ejemplo, un supuesto es que los depredadores tienen una respuesta funcional lineal a las presas: la tasa de muertes aumenta en proporción a la tasa de encuentros. Si esta tasa está limitada por el tiempo dedicado a manipular cada captura, entonces las poblaciones de presas pueden alcanzar densidades por encima de las cuales los depredadores no pueden controlarlas. [144] Otro supuesto es que todos los individuos presa son idénticos. En realidad, los depredadores tienden a seleccionar individuos jóvenes, débiles y enfermos, lo que permite que las poblaciones de presas vuelvan a crecer. [147]
Muchos factores pueden estabilizar las poblaciones de depredadores y presas. [148] Un ejemplo es la presencia de múltiples depredadores, particularmente generalistas que se sienten atraídos por una especie de presa dada si es abundante y buscan en otro lugar si no lo es. [149] Como resultado, los ciclos poblacionales tienden a encontrarse en ecosistemas templados y subárticos del norte porque las redes alimentarias son más simples. [150] El sistema liebre de raquetas de nieve-lince es subártico, pero incluso esto involucra a otros depredadores, incluidos coyotes, azores y búhos cornudos , y el ciclo se refuerza por las variaciones en el alimento disponible para las liebres. [151]
Se han desarrollado una serie de modelos matemáticos flexibilizando las suposiciones realizadas en el modelo de Lotka-Volterra; estos permiten que los animales tengan distribuciones geográficas o migren ; que tengan diferencias entre individuos, como sexos y una estructura de edad , de modo que solo algunos individuos se reproduzcan; que vivan en un entorno variable, como con estaciones cambiantes ; [152] [153] y que analicen las interacciones de más de dos especies a la vez. Estos modelos predicen dinámicas de población depredador-presa muy diferentes y a menudo caóticas . [152] [154] La presencia de áreas de refugio , donde las presas están a salvo de los depredadores, puede permitir que las presas mantengan poblaciones más grandes, pero también puede desestabilizar la dinámica. [155] [156] [157] [158]
La depredación data de antes del surgimiento de los carnívoros comúnmente reconocidos por cientos de millones (quizás miles de millones) de años. La depredación ha evolucionado repetidamente en diferentes grupos de organismos. [5] [159] El surgimiento de las células eucariotas alrededor de 2,7 millones de años atrás, el surgimiento de los organismos multicelulares alrededor de 2 millones de años atrás y el surgimiento de los depredadores móviles (alrededor de 600 millones de años - 2 millones de años, probablemente alrededor de 1 millón de años) se han atribuido a un comportamiento depredador temprano, y muchos restos muy tempranos muestran evidencia de pozos u otras marcas atribuidas a pequeñas especies de depredadores. [5] Probablemente desencadenó importantes transiciones evolutivas, incluida la llegada de las células , los eucariotas , la reproducción sexual , la multicelularidad , el aumento del tamaño, la movilidad (incluido el vuelo de los insectos [160] ) y los caparazones y exoesqueletos blindados. [5]
Los primeros depredadores fueron organismos microbianos que engullían o pastaban a otros. Debido a que el registro fósil es pobre, estos primeros depredadores podrían remontarse a entre 1 y más de 2,7 mil millones de años atrás. [5] La depredación se volvió visiblemente importante poco antes del período Cámbrico , hace unos 550 millones de años , como lo demuestra el desarrollo casi simultáneo de la calcificación en animales y algas, [161] y la excavación para evitar la depredación . Sin embargo, los depredadores habían estado pastando microorganismos desde hace al menos 1000 millones de años , [5] [162] [163] con evidencia de depredación selectiva (en lugar de aleatoria) en una época similar. [164]
Auroralumina attenboroughii es un cnidario del grupo corona del Ediacárico(557–562 millones de años atrás, unos 20 millones de años antes de la explosión cámbrica) del bosque de Charnwood , Inglaterra. Se cree que es uno de los primeros animales depredadores, que capturaba presas pequeñas con sus nematocistos , como lo hacen los cnidarios modernos. [165]
El registro fósil demuestra una larga historia de interacciones entre depredadores y sus presas desde el período Cámbrico en adelante, mostrando por ejemplo que algunos depredadores perforaban las conchas de moluscos bivalvos y gasterópodos , mientras que otros comían estos organismos rompiendo sus conchas. [166] Entre los depredadores cámbricos había invertebrados como los anomalocarídidos con apéndices adecuados para agarrar presas, grandes ojos compuestos y mandíbulas hechas de un material duro como el del exoesqueleto de un insecto. [167] Algunos de los primeros peces en tener mandíbulas fueron los placodermos acorazados y principalmente depredadores de los períodos Silúrico al Devónico , uno de los cuales, el Dunkleosteus de 6 m (20 pies) , se considera el primer "superdepredador" vertebrado del mundo , que se alimenta de otros depredadores. [168] [169] Los insectos desarrollaron la capacidad de volar en el Carbonífero Temprano o el Devónico Tardío, lo que les permitió, entre otras cosas, escapar de los depredadores. [160] Entre los depredadores más grandes que han vivido se encuentran los dinosaurios terópodos como el Tyrannosaurus del período Cretácico . Se alimentaban de dinosaurios herbívoros como los hadrosaurios , los ceratopsianos y los anquilosaurios . [170]


.png/1280px-Dunkleosteus_terrelli_(2024).png)

Los humanos, como omnívoros , son hasta cierto punto depredadores, [171] usando armas y herramientas para pescar , [172] cazar y atrapar animales. [173] También utilizan otras especies depredadoras como perros , cormoranes , [174] y halcones para atrapar presas para comer o por deporte. [175] Dos depredadores de tamaño mediano, perros y gatos, son los animales que se mantienen con más frecuencia como mascotas en las sociedades occidentales. [176] [177] Los cazadores humanos, incluidos los san del sur de África, utilizan la caza de persistencia , una forma de depredación por persecución en la que el perseguidor puede ser más lento que la presa, como un antílope kudu en distancias cortas, pero lo sigue en el calor del mediodía hasta que se agota, una persecución que puede durar hasta cinco horas. [178] [179]
En el control biológico de plagas , se introducen depredadores (y parasitoides) del área de distribución natural de una plaga para controlar las poblaciones, con el riesgo de causar problemas imprevistos. Los depredadores naturales, siempre que no dañen a las especies que no son plagas, son una forma ecológica y sostenible de reducir los daños a los cultivos y una alternativa al uso de agentes químicos como los pesticidas . [180]

En el cine, la idea del depredador como un enemigo humanoide peligroso se utiliza en la película de acción y terror de ciencia ficción de 1987 Depredador y sus tres secuelas . [181] [182] Un depredador aterrador, un gigantesco tiburón blanco devorador de hombres , también es central en el thriller de Steven Spielberg de 1974, Tiburón . [183]
Entre la poesía sobre el tema de la depredación, se puede explorar la conciencia de un depredador, como en Pike de Ted Hughes . [184] La frase "Naturaleza, roja en dientes y garras" del poema de Alfred, Lord Tennyson de 1849 " In Memoriam AHH " ha sido interpretada como una referencia a la lucha entre depredadores y presas. [185]
En la mitología y las fábulas populares, los depredadores como el zorro y el lobo tienen reputaciones mixtas. [186] El zorro era un símbolo de fertilidad en la antigua Grecia, pero un demonio del clima en el norte de Europa y una criatura del diablo en el cristianismo primitivo; el zorro se presenta como astuto, codicioso y astuto en las fábulas desde Esopo en adelante. [186] El lobo feroz es conocido por los niños en cuentos como Caperucita Roja , pero es una figura demoníaca en las sagas islandesas de Edda , donde el lobo Fenrir aparece en el final apocalíptico del mundo . [186] En la Edad Media, se extendió la creencia en los hombres lobo , hombres transformados en lobos. [186] En la antigua Roma y en el antiguo Egipto, se adoraba al lobo, y la loba aparece en el mito fundador de Roma, amamantando a Rómulo y Remo . [186] Más recientemente, en El libro de la selva de Rudyard Kipling de 1894 , Mowgli es criado por la manada de lobos. [186] Las actitudes hacia los grandes depredadores en América del Norte, como el lobo, el oso pardo y el puma, han cambiado de hostilidad o ambivalencia, acompañada de persecución activa, hacia actitudes positivas y protectoras en la segunda mitad del siglo XX. [187]
{{cite journal}}: Requiere citar revista |journal=( ayuda )Cursorial hunting strategies range from one extreme of transient acceleration, power and speed to the other extreme of persistence and endurance with prey being fatigued to facilitate capture.Dogs and humans are considered to rely on endurance rather than outright speed and manoeuvrability for success when hunting cursorially.
{{cite book}}: CS1 maint: multiple names: authors list (link)Hughes's earliest books contained a bewildering profusion of poems between their covers: ... fish and fowl, beasts of the field and forest, vigorous embodiments of predators and prey. Hughes as a student had taken up anthropology, not literature, and he chose to meditate his way into trancelike states of preconsciousness before committing poems to paper. His poems, early or late, enter into the relations of living creatures; they move in close to animal consciousness: The Thought-Fox, Esther's Tomcat, Pike.


.jpg/1280px-Hawk_eating_prey_(cropped).jpg)

![Auroralumina attenboroughii, un depredador ediacárico (hace unos 560 millones de años). Era un cnidario del grupo troncal que capturaba presas con sus nematocistos.[165]](http://upload.wikimedia.org/wikipedia/commons/thumb/6/63/Auroralumina_attenboroughii_reconstruction.jpg/1280px-Auroralumina_attenboroughii_reconstruction.jpg)

