La progesterona y algunos de sus metabolitos, como la 5β-dihidroprogesterona , son agonistas del receptor X de pregnano (PXR), [30] aunque débilmente ( EC 50 >10 μM). [31] De acuerdo con esto, la progesterona induce varias enzimas hepáticas del citocromo P450 , [32] como CYP3A4 , [33] [34] especialmente durante el embarazo cuando las concentraciones son mucho más altas de lo habitual. [35] Se ha descubierto que las mujeres perimenopáusicas tienen una mayor actividad de CYP3A4 en relación con los hombres y las mujeres posmenopáusicas, y se ha inferido que esto puede deberse a los niveles más altos de progesterona presentes en las mujeres perimenopáusicas. [33]
La progesterona modula la actividad de los canales de Ca 2+ dependientes del voltaje de los canales de cationes de los espermatozoides ( CatSper ) . Dado que los óvulos liberan progesterona, los espermatozoides pueden usarla como señal de retorno para nadar hacia los óvulos ( quimiotaxis ). Como resultado, se ha sugerido que las sustancias que bloquean el sitio de unión de la progesterona en los canales de CatSper podrían potencialmente usarse en la anticoncepción masculina . [36] [37]
Función biológica
Durante el ciclo menstrual, los niveles de estradiol (un estrógeno) varían en un 200 por ciento. Los niveles de progesterona varían en más del 1200 por ciento. [38]
Interacciones hormonales
La progesterona tiene una serie de efectos fisiológicos que se amplifican en presencia de estrógenos . Los estrógenos a través de los receptores de estrógeno (RE) inducen o regulan positivamente la expresión del PR. [39] Un ejemplo de esto es en el tejido mamario , donde los estrógenos permiten que la progesterona media el desarrollo lobuloalveolar . [40] [41] [42]
Los niveles elevados de progesterona reducen de forma potente la actividad de retención de sodio de la aldosterona, lo que produce natriuresis y una reducción del volumen de líquido extracelular. Por otra parte, la retirada de progesterona se asocia a un aumento temporal de la retención de sodio (natriuresis reducida, con un aumento del volumen de líquido extracelular) debido al aumento compensatorio de la producción de aldosterona, que combate el bloqueo del receptor de mineralocorticoides por el nivel previamente elevado de progesterona. [43]
Diferenciación sexual temprana
La progesterona desempeña un papel en la diferenciación sexual humana temprana. [44] La progesterona placentaria es la materia prima para la 5α-dihidrotestosterona (DHT) producida a través de la vía de puerta trasera que opera en múltiples tejidos no gonadales del feto , [45] mientras que las deficiencias en esta vía conducen a la subvirilización del feto masculino, lo que resulta en un desarrollo incompleto de los genitales masculinos. [46] [47] La DHT es un potente andrógeno que es responsable del desarrollo de los genitales masculinos, incluido el pene y el escroto .
Durante el desarrollo fetal temprano, las gónadas indiferenciadas pueden convertirse en testículos u ovarios. La presencia del cromosoma Y conduce al desarrollo de los testículos. Los testículos luego producen testosterona, que se convierte en DHT a través de la enzima 5α-reductasa . La DHT es un potente andrógeno que es responsable de la masculinización de los genitales externos y el desarrollo de la glándula prostática. La progesterona, producida por la placenta durante el embarazo, desempeña un papel en la diferenciación sexual fetal al servir como molécula precursora para la síntesis de DHT a través de la vía de la puerta trasera. En ausencia de niveles adecuados de enzimas esteroidogénicas durante el desarrollo fetal, la vía de la puerta trasera para la síntesis de DHT puede volverse deficiente, lo que lleva a la submasculinización del feto masculino. Esto puede resultar en el desarrollo de genitales ambiguos o incluso genitales femeninos en algunos casos. Por lo tanto, tanto la DHT como la progesterona desempeñan un papel crucial en la diferenciación sexual fetal temprana, y la progesterona actúa como una molécula precursora para la síntesis de DHT y la DHT promueve el desarrollo de los genitales masculinos. [44]
La progesterona tiene efectos clave a través de la señalización no genómica en los espermatozoides humanos a medida que migran a través del tracto reproductivo femenino antes de que se produzca la fertilización , aunque el receptor o los receptores aún no se han identificado. [48] La caracterización detallada de los eventos que ocurren en los espermatozoides en respuesta a la progesterona ha dilucidado ciertos eventos, incluidos los transitorios de calcio intracelular y los cambios mantenidos, [49] oscilaciones lentas de calcio, [50] que ahora se cree que posiblemente regulan la motilidad. [51] Es producida por los ovarios. [52] También se ha demostrado que la progesterona tiene efectos sobre los espermatozoides de pulpo. [53]
A la progesterona a veces se la denomina la "hormona del embarazo ", [54] y tiene muchas funciones relacionadas con el desarrollo del feto:
Durante la implantación y la gestación , la progesterona parece disminuir la respuesta inmune materna para permitir la aceptación del embarazo. [56]
La progesterona disminuye la contractilidad del músculo liso uterino . [54] Este efecto contribuye a la prevención del parto prematuro . [56] Los estudios han demostrado que en las mujeres embarazadas de un solo feto, asintomáticas en la etapa prenatal y con un alto riesgo de dar a luz prematuramente de forma espontánea, se ha descubierto que la medicación vaginal con progesterona es eficaz para prevenir el parto prematuro espontáneo. Las personas que tienen un alto riesgo de dar a luz prematuramente de forma espontánea son aquellas que tienen un cuello uterino corto de menos de 25 mm o que han dado a luz prematuramente de forma espontánea anteriormente. Aunque generalmente se considera que los partos prematuros son de menos de 37 semanas, estos estudios encontraron que la progesterona vaginal se asocia con menos partos prematuros de menos de 34 semanas. [57]
Una caída en los niveles de progesterona es posiblemente un paso que facilita el inicio del parto .
Además, la progesterona inhibe la lactancia durante el embarazo. La caída de los niveles de progesterona después del parto es uno de los desencadenantes de la producción de leche.
La progesterona desempeña un papel importante en el desarrollo de los senos . Junto con la prolactina , media la maduración lobuloalveolar de las glándulas mamarias durante el embarazo para permitir la producción de leche y, por lo tanto, la lactancia y el amamantamiento de la descendencia después del parto (nacimiento). [58] El estrógeno induce la expresión de PR en el tejido mamario y, por lo tanto, la progesterona depende del estrógeno para mediar el desarrollo lobuloalveolar. [40] [41] [42] Se ha descubierto que RANKLActivador del receptor del ligando del factor nuclear kappa-Bes un mediador crítico de la maduración lobuloalveolar inducida por progesterona. [59] Los ratones knock out de RANKL muestran un fenotipo mamario casi idéntico al de los ratones knock out de PR, incluido el desarrollo ductal mamario normal pero un fracaso completo del desarrollo de las estructuras lobuloalveolares. [59]
Desarrollo ductal
Aunque en un grado mucho menor que el estrógeno, que es el principal mediador del desarrollo ductal mamario (a través del ERα ), [60] [61] la progesterona también puede estar involucrada en el desarrollo ductal de las glándulas mamarias hasta cierto punto. [62] Los ratones knock out de PR o los ratones tratados con el antagonista de PR mifepristona muestran un desarrollo ductal mamario retardado aunque por lo demás normal en la pubertad. [62] Además, los ratones modificados para tener sobreexpresión de PRA muestran hiperplasia ductal, [59] y la progesterona induce el crecimiento ductal en la glándula mamaria del ratón. [62] La progesterona media el desarrollo ductal principalmente a través de la inducción de la expresión de anfiregulina , el mismo factor de crecimiento cuya expresión el estrógeno induce principalmente para mediar el desarrollo ductal. [62] Estos hallazgos en animales sugieren que, si bien no es esencial para el desarrollo completo del conducto mamario, la progesterona parece desempeñar un papel potenciador o acelerador del desarrollo del conducto mamario mediado por estrógenos. [62]
Riesgo de cáncer de mama
La progesterona también parece estar involucrada en la fisiopatología del cáncer de mama , aunque su papel, y si es un promotor o inhibidor del riesgo de cáncer de mama, no se ha dilucidado por completo. [63] [64] Se ha descubierto que la mayoría de las progestinas , o progestágenos sintéticos , como el acetato de medroxiprogesterona , aumentan el riesgo de cáncer de mama en personas posmenopáusicas en combinación con estrógeno como componente de la terapia hormonal menopáusica . [65] [64] La combinación de progesterona oral natural o la progestina atípica didrogesterona con estrógeno se ha asociado con un menor riesgo de cáncer de mama que las progestinas más estrógeno. [66] [67] [68] Sin embargo, esto puede ser simplemente un artefacto de los bajos niveles de progesterona producidos con progesterona oral. [63] [69] Se necesita más investigación sobre el papel de la progesterona en el cáncer de mama. [64]
La terapia de reemplazo hormonal, que consiste en un tratamiento sistémico con estrógeno solo o en combinación con un progestágeno, tiene efectos beneficiosos considerables y bien documentados sobre la piel de las personas posmenopáusicas. [70] [71] Estos beneficios incluyen un mayor contenido de colágeno de la piel, grosor y elasticidad de la piel e hidratación de la piel y lípidos superficiales. [70] [71] Se ha descubierto que el estrógeno tópico tiene efectos beneficiosos similares sobre la piel. [70] Además, un estudio ha descubierto que la crema tópica de progesterona al 2% aumenta significativamente la elasticidad y firmeza de la piel y disminuye observablemente las arrugas en personas peri y posmenopáusicas. [71] La hidratación de la piel y los lípidos superficiales, por otro lado, no cambiaron significativamente con la progesterona tópica. [71]
Estos hallazgos sugieren que la progesterona, al igual que el estrógeno, también tiene efectos beneficiosos sobre la piel y puede ser un protector independiente contra el envejecimiento de la piel. [71]
La Dra. Diana Fleischman , de la Universidad de Portsmouth , y sus colegas buscaron una relación entre la progesterona y las actitudes sexuales en 92 mujeres. Su investigación, publicada en Archives of Sexual Behavior, descubrió que las mujeres que tenían niveles más altos de progesterona obtuvieron puntuaciones más altas en un cuestionario que medía la motivación homoerótica. También descubrieron que los hombres que tenían niveles altos de progesterona tenían más probabilidades de tener puntuaciones más altas de motivación homoerótica después de la preparación afiliativa en comparación con los hombres con niveles bajos de progesterona. [73] [74] [75] [76]
Los neuroesteroides son neuromoduladores , neuroprotectores , neurogénicos y regulan la neurotransmisión y la mielinización . [78] Los efectos de la progesterona como neuroesteroide están mediados predominantemente a través de sus interacciones con PR no nucleares, a saber, los mPR y PGRMC1 , así como ciertos otros receptores, como los receptores σ 1 y nACh. [79]
Daño cerebral
Estudios previos han demostrado que la progesterona favorece el desarrollo normal de las neuronas en el cerebro y que la hormona tiene un efecto protector sobre el tejido cerebral dañado. Se ha observado en modelos animales que las hembras tienen una menor susceptibilidad a las lesiones cerebrales traumáticas y se ha planteado la hipótesis de que este efecto protector se debe al aumento de los niveles circulantes de estrógeno y progesterona en las hembras. [80]
Mecanismo propuesto
El mecanismo de los efectos protectores de la progesterona puede ser la reducción de la inflamación que sigue a un traumatismo y una hemorragia cerebral. [81] [82]
Se cree que el daño causado por una lesión cerebral traumática es causado en parte por la despolarización masiva que conduce a la excitotoxicidad . Una forma en que la progesterona ayuda a aliviar parte de esta excitotoxicidad es bloqueando los canales de calcio dependientes del voltaje que desencadenan la liberación de neurotransmisores . [83] Lo hace manipulando las vías de señalización de los factores de transcripción involucrados en esta liberación. Otro método para reducir la excitotoxicidad es mediante la regulación positiva del GABA A , un receptor inhibidor generalizado de neurotransmisores. [84]
También se ha demostrado que la progesterona previene la apoptosis en las neuronas, una consecuencia común de las lesiones cerebrales. [85] Lo hace inhibiendo las enzimas implicadas en la vía de la apoptosis específicamente en relación con las mitocondrias, como la caspasa 3 activada y el citocromo c .
La progesterona no solo ayuda a prevenir más daños, sino que también se ha demostrado que ayuda en la neuroregeneración . [86] Uno de los efectos graves de la lesión cerebral traumática incluye el edema. Los estudios en animales muestran que el tratamiento con progesterona conduce a una disminución en los niveles de edema al aumentar la concentración de macrófagos y microglia enviada al tejido lesionado. [83] [87] Esto se observó en forma de reducción de fugas de la barrera hematoencefálica en la recuperación secundaria en ratas tratadas con progesterona. Además, se observó que la progesterona tiene propiedades antioxidantes , reduciendo la concentración de radicales libres de oxígeno más rápido que sin ella. [84] También hay evidencia de que la adición de progesterona también puede ayudar a remielinizar los axones dañados debido al trauma, restaurando algo de conducción de señales neuronales perdidas. [84] Otra forma en que la progesterona ayuda en la regeneración incluye aumentar la circulación de células progenitoras endoteliales en el cerebro. [88] Esto ayuda a que crezca nueva vasculatura alrededor del tejido cicatricial, lo que ayuda a reparar el área de la lesión.
Adicción
La progesterona mejora la función de los receptores de serotonina en el cerebro, por lo que un exceso o déficit de progesterona tiene el potencial de provocar problemas neuroquímicos importantes. Esto proporciona una explicación de por qué algunas personas recurren a sustancias que mejoran la actividad de la serotonina , como la nicotina , el alcohol y el cannabis , cuando sus niveles de progesterona caen por debajo de los niveles óptimos. [89]
Las diferencias de sexo en los niveles hormonales pueden inducir a las mujeres a responder de manera diferente que los hombres a la nicotina. Cuando las mujeres atraviesan cambios cíclicos o diferentes fases de transición hormonal (menopausia, embarazo, adolescencia), se producen cambios en sus niveles de progesterona. [90] Por lo tanto, las mujeres tienen una mayor vulnerabilidad biológica a los efectos de refuerzo de la nicotina en comparación con los hombres y la progesterona puede utilizarse para contrarrestar esta mayor vulnerabilidad. Esta información apoya la idea de que la progesterona puede afectar el comportamiento. [89]
Al igual que la nicotina, la cocaína también aumenta la liberación de dopamina en el cerebro. Este neurotransmisor está involucrado en el centro de recompensa y es uno de los principales neurotransmisores relacionados con el abuso y la dependencia de sustancias. En un estudio sobre consumidores de cocaína, se informó que la progesterona reducía el ansia y la sensación de ser estimulado por la cocaína. Por lo tanto, se sugirió que la progesterona era un agente que disminuye el ansia de cocaína al reducir las propiedades dopaminérgicas de la droga. [91]
Societal
En un estudio de 2012 de la Universidad de Ámsterdam sobre 120 mujeres, la fase lútea de las mujeres (niveles más altos de progesterona y niveles crecientes de estrógeno) se correlacionó con un menor nivel de comportamiento competitivo en escenarios de juegos de azar y concursos de matemáticas, mientras que su fase premenstrual (niveles de progesterona en marcada disminución y niveles decrecientes de estrógeno) se correlacionó con un mayor nivel de comportamiento competitivo. [92]
Otros efectos
La progesterona también tiene un papel en la elasticidad de la piel y la fortaleza de los huesos, en la respiración , en el tejido nervioso y en la sexualidad femenina , y la presencia de receptores de progesterona en ciertos tejidos musculares y grasos puede indicar un papel en las proporciones sexualmente dimórficas de aquellos. [93] [ ¿ enlace infractor? ]
Se dice que durante el embarazo, la progesterona disminuye la irritabilidad uterina. [94]
Durante el embarazo, la progesterona ayuda a suprimir las respuestas inmunes de la madre a los antígenos fetales, lo que evita el rechazo del feto. [94]
La progesterona normaliza la coagulación sanguínea y el tono vascular, los niveles de zinc y cobre , los niveles de oxígeno celular y el uso de las reservas de grasa para obtener energía.
La progesterona puede afectar la salud de las encías, aumentando el riesgo de gingivitis (inflamación de las encías). [99]
La progesterona parece prevenir el cáncer de endometrio (que afecta el revestimiento del útero) al regular los efectos del estrógeno.
La progesterona desempeña un papel importante en la señalización de la liberación de insulina y la función pancreática, y puede afectar la susceptibilidad a la diabetes o la diabetes gestacional. [100] [101]
Se encontró que los niveles de progesterona en la sangre eran más bajos en aquellas que tenían mayor peso y un IMC más alto entre aquellas que quedaron embarazadas mediante fertilización in vitro. [102]
Los datos actuales muestran que la progesterona micronizada, que es químicamente idéntica a la progesterona producida en el cuerpo humano, en combinación con estrógenos en la terapia hormonal para la menopausia no parece tener efectos significativos sobre la tromboembolia venosa (coágulos de sangre en las venas) y el accidente cerebrovascular isquémico (falta de flujo sanguíneo al cerebro debido al bloqueo de un vaso sanguíneo que lo irriga). Sin embargo, es necesario realizar más estudios para ver si la progesterona micronizada sola o en combinación con la terapia hormonal para la menopausia modifica el riesgo de infartos de miocardio (ataques cardíacos). [103]
Aún no se han realizado estudios sobre los efectos de la progesterona micronizada en la caída del cabello debido a la menopausia. [104]
A pesar de las sugerencias de utilizar terapia hormonal para prevenir la pérdida de masa muscular en personas posmenopáusicas (de 50 años o más), no se ha demostrado que la terapia hormonal menopáusica que involucra estrógeno solo o estrógeno y progesterona preserve la masa muscular. [105] La terapia hormonal menopáusica tampoco produce una reducción del peso corporal, una reducción del IMC o un cambio en el metabolismo de la glucosa. [106]
Bioquímica
Biosíntesis
Esteroidogénesis , que muestra la progesterona entre los progestágenos en el área amarilla. [107]
El colesterol sufre una doble oxidación para producir 22 R -hidroxicolesterol y luego 20α,22 R -dihidroxicolesterol . Este diol vecinal se oxida aún más con pérdida de la cadena lateral que comienza en la posición C22 para producir pregnenolona. Esta reacción es catalizada por el citocromo P450scc .
La pregnenolona y la progesterona también pueden ser sintetizadas por la levadura . [109]
Los ovarios secretan aproximadamente 25 mg de progesterona por día, mientras que las glándulas suprarrenales producen alrededor de 2 mg de progesterona por día. [110]
El metabolismo de la progesterona es rápido y extenso y ocurre principalmente en el hígado , [112] [113] [114] aunque las enzimas que metabolizan la progesterona también se expresan ampliamente en el cerebro , la piel y varios otros tejidos extrahepáticos . [77] [115] La progesterona tiene una vida media de eliminación de solo aproximadamente 5 minutos en circulación . [112] El metabolismo de la progesterona es complejo y puede formar hasta 35 metabolitos no conjugados diferentes cuando se ingiere por vía oral. [114] [116] La progesterona es altamente susceptible a la reducción enzimática a través de reductasas e hidroxiesteroide deshidrogenasas debido a su doble enlace (entre las posiciones C4 y C5) y sus dos cetonas (en las posiciones C3 y C20). [114]
Porciones relativamente pequeñas de progesterona se hidroxilan a través de la 17α-hidroxilasa (CYP17A1) y la 21-hidroxilasa (CYP21A2) en 17α-hidroxiprogesterona y 11-desoxicorticosterona (21-hidroxiprogesterona), respectivamente, [116] y los pregnanetrioles se forman secundariamente a la 17α-hidroxilación. [123] [124] Cantidades incluso más pequeñas de progesterona también pueden hidroxilarse a través de la 11β-hidroxilasa (CYP11B1) y en menor medida a través de la aldosterona sintasa (CYP11B2) en 11β-hidroxiprogesterona . [125] [126] [44] Además, la progesterona puede hidroxilarse en el hígado por otras enzimas del citocromo P450 que no son específicas de los esteroides. [127] La 6β-hidroxilación, que es catalizada principalmente por CYP3A4 , es la principal transformación y es responsable de aproximadamente el 70% del metabolismo de la progesterona mediado por el citocromo P450. [127] Otras vías incluyen la 6α-, 16α- y 16β-hidroxilación. [114] Sin embargo, el tratamiento de mujeres con ketoconazol , un potente inhibidor del CYP3A4, tuvo efectos mínimos en los niveles de progesterona, produciendo solo un aumento leve y no significativo, y esto sugiere que las enzimas del citocromo P450 juegan solo un papel pequeño en el metabolismo de la progesterona. [128]
Niveles
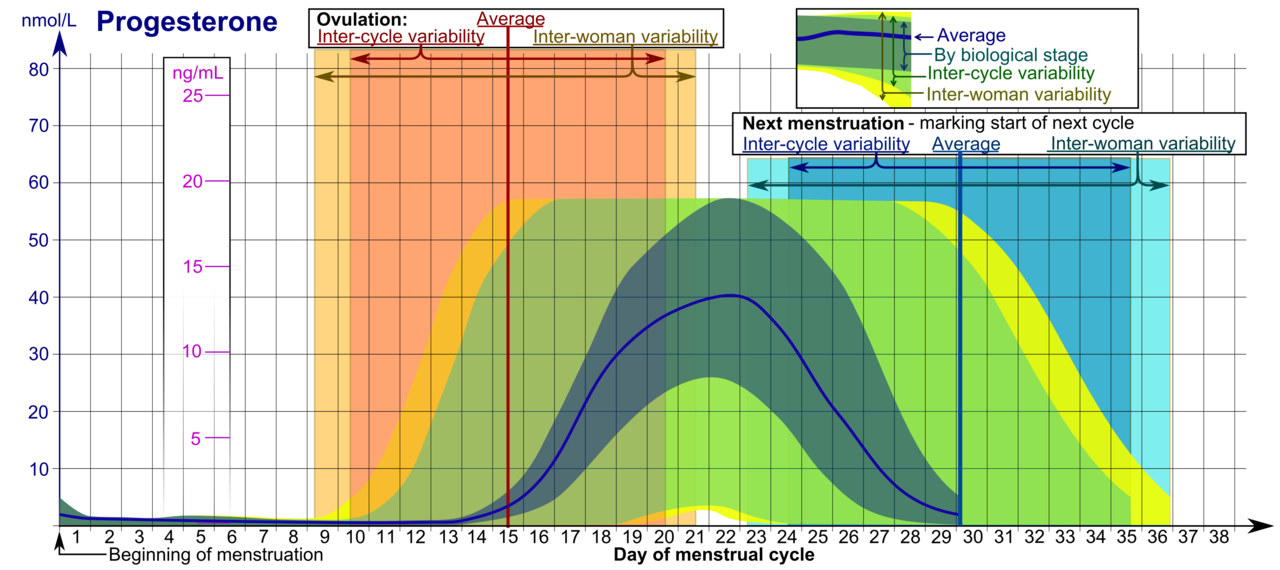
Niveles de progesterona a lo largo del ciclo menstrual en mujeres con ciclos normales y ovulatorias. [130] Las líneas horizontales son los niveles integrados medios para cada curva. La línea vertical es la mitad del ciclo.
Los niveles de progesterona son relativamente bajos durante la fase preovulatoria del ciclo menstrual , aumentan después de la ovulación y se elevan durante la fase lútea , como se muestra en el diagrama anterior. Los niveles de progesterona tienden a ser inferiores a 2 ng/mL antes de la ovulación y superiores a 5 ng/mL después de la ovulación. Si se produce un embarazo , se libera gonadotropina coriónica humana , lo que mantiene el cuerpo lúteo y le permite mantener los niveles de progesterona. Entre las semanas 7 y 9, la placenta comienza a producir progesterona en lugar del cuerpo lúteo en un proceso llamado desplazamiento lúteo-placentario. [131]
Después del cambio de fase lútea a placentaria, los niveles de progesterona comienzan a aumentar aún más y pueden alcanzar entre 100 y 200 ng/mL al término del embarazo. Se ha discutido si una disminución de los niveles de progesterona es crítica para el inicio del parto y puede ser específica de cada especie. Después de la expulsión de la placenta y durante la lactancia, los niveles de progesterona son muy bajos.
Los niveles de progesterona son bajos en niños y personas posmenopáusicas. [132] Los hombres adultos tienen niveles similares a los de las mujeres durante la fase folicular del ciclo menstrual.
Rangos
Los resultados de los análisis de sangre siempre deben interpretarse utilizando los rangos de referencia proporcionados por el laboratorio que realizó los resultados. A continuación se enumeran algunos ejemplos de rangos de referencia.
Rangos de referencia para el contenido de progesterona en sangre durante el ciclo menstrual
Niveles de progesterona durante el ciclo menstrual . [143] • Los rangos denotados Por etapa biológica pueden usarse en ciclos menstruales monitoreados de cerca con respecto a otros marcadores de su progresión biológica, con la escala de tiempo comprimida o estirada a cuánto más rápido o más lento, respectivamente, progresa el ciclo en comparación con un ciclo promedio. • Los rangos denotados Variabilidad interciclo son más apropiados para usar en ciclos no monitoreados con solo el comienzo de la menstruación conocido, pero donde la mujer conoce con precisión las duraciones promedio de su ciclo y el momento de la ovulación, y que son algo regulares en promedio, con la escala de tiempo comprimida o estirada a cuánto es más corta o más larga la duración promedio del ciclo de una mujer, respectivamente, que el promedio de la población. • Los rangos denotados Variabilidad intermujer son más apropiados para usar cuando se desconocen las duraciones promedio del ciclo y el momento de la ovulación, pero solo se da el comienzo de la menstruación.
Durante el embarazo humano, los ovarios y la placenta producen progesterona en cantidades cada vez mayores . Al principio, la fuente es el cuerpo lúteo que ha sido "rescatado" por la presencia de gonadotropina coriónica humana (hCG) del feto. Sin embargo, después de la octava semana, la producción de progesterona se desplaza a la placenta. La placenta utiliza el colesterol materno como sustrato inicial, y la mayor parte de la progesterona producida ingresa a la circulación materna, pero una parte es captada por la circulación fetal y utilizada como sustrato para los corticosteroides fetales. A término, la placenta produce alrededor de 250 mg de progesterona por día.
Otra fuente animal de progesterona son los productos lácteos. Después del consumo de productos lácteos, el nivel de progesterona biodisponible aumenta. [144]
Plantas
En al menos una planta, Juglans regia , se ha detectado progesterona. [145] Además, se encuentran esteroides similares a la progesterona en Dioscorea mexicana . Dioscorea mexicana es una planta que forma parte de la familia del ñame originaria de México . [146] Contiene un esteroide llamado diosgenina que se toma de la planta y se convierte en progesterona. [147] La diosgenina y la progesterona también se encuentran en otras especies de Dioscorea , así como en otras plantas que no están estrechamente relacionadas, como el fenogreco .
Otra planta que contiene sustancias fácilmente convertibles en progesterona es la Dioscorea pseudojaponica , originaria de Taiwán . Las investigaciones han demostrado que el ñame taiwanés contiene saponinas , esteroides que pueden convertirse en diosgenina y, de ahí, en progesterona. [148]
La progesterona se utiliza como medicamento . Se utiliza en combinación con estrógenos principalmente en la terapia hormonal para los síntomas de la menopausia y los niveles bajos de hormonas sexuales . [116] [152] También se puede utilizar sola para tratar los síntomas de la menopausia. Los estudios han demostrado que la progesterona transdérmica (parche cutáneo) y la progesterona micronizada oral son tratamientos efectivos para ciertos síntomas de la menopausia, como los sofocos y los sudores nocturnos, que también se conocen como síntomas vasomotores o VMS. [153]
También se utiliza para apoyar el embarazo y la fertilidad y para tratar trastornos ginecológicos . [154] [155] [156] [157] Se ha demostrado que la progesterona previene el aborto espontáneo en aquellas con 1) sangrado vaginal temprano en su embarazo actual y 2) antecedentes de aborto espontáneo. [158] La progesterona se puede tomar por vía oral , a través de la vagina y mediante inyección en el músculo o la grasa , entre otras vías . [116]
Química
Una muestra de progesterona
La progesterona es un esteroide pregnano natural y también se conoce como pregn-4-eno-3,20-diona. [159] [160] Tiene un doble enlace ( 4-eno ) entre las posiciones C4 y C5 y dos grupos cetona (3,20- diona ), uno en la posición C3 y el otro en la posición C20. [159] [160]
Síntesis
La progesterona se produce comercialmente por semisíntesis. Se utilizan dos rutas principales: una a partir de la diosgenina del ñame, desarrollada por primera vez por Marker en 1940, y otra basada en fitoesteroles de soja , que se amplió en la década de 1970. También se han descrito otras semisíntesis de progesterona (no necesariamente económicas) a partir de una variedad de esteroides. Por ejemplo, la cortisona se puede desoxigenar simultáneamente en la posición C-17 y C-21 mediante el tratamiento con yodotrimetilsilano en cloroformo para producir 11-ceto-progesterona (ketogestina), que a su vez se puede reducir en la posición 11 para producir progesterona. [161]
Semisíntesis de progesterona a partir de diosgenina . [ 162]
El intermediario 16-DPA es importante para la síntesis de muchos otros esteroides de importancia médica. Un enfoque muy similar puede producir 16-DPA a partir de solanina . [163]
En 1971, WS Johnson informó de una síntesis total de progesterona . [169] La síntesis comienza con la reacción de la sal de fosfonio 7 con fenil litio para producir el iluro de fosfonio 8. El iluro 8 se hace reaccionar con un aldehído para producir el alqueno 9. Los grupos protectores de cetal de 9 se hidrolizan para producir la dicetona 10 , que a su vez se cicla para formar la ciclopentenona 11. La cetona de 11 se hace reaccionar con metil litio para producir el alcohol terciario 12 , que a su vez se trata con ácido para producir el catión terciario 13. El paso clave de la síntesis es la ciclización del catión π de 13 en el que los anillos B, C y D del esteroide se forman simultáneamente para producir 14 . Este paso se asemeja a la reacción de ciclización catiónica utilizada en la biosíntesis de esteroides y, por lo tanto, se denomina biomimético . En el siguiente paso, el ortoéster de enol se hidroliza para producir la cetona 15. Luego, el anillo A del ciclopenteno se abre mediante oxidación con ozono para producir 16. Finalmente, la dicetona 17 sufre una condensación aldólica intramolecular mediante tratamiento con hidróxido de potasio acuoso para producir progesterona. [169]
Historia
George W. Corner y Willard M. Allen descubrieron la acción hormonal de la progesterona en 1929. [17] [170] [171] [172] Entre 1931 y 1932, se había aislado material cristalino casi puro de alta actividad progestacional del cuerpo lúteo de animales, y en 1934, se había refinado y obtenido progesterona cristalina pura y se había determinado la estructura química de la progesterona. [17] [171] Esto lo logró Adolf Butenandt en el Instituto Químico de la Universidad Técnica de Danzig , quien extrajo este nuevo compuesto de varios miles de litros de orina . [173]
La síntesis química de progesterona a partir de estigmasterol y pregnanediol se logró más tarde ese año. [171] [174] Hasta este punto, la progesterona, conocida genéricamente como hormona del cuerpo lúteo, había sido referida por varios grupos con diferentes nombres, incluyendo corporina, luteína, luteosterona y progestina. [17] [175] En 1935, en el momento de la Segunda Conferencia Internacional sobre la Estandarización de las Hormonas Sexuales en Londres, Inglaterra , se llegó a un compromiso entre los grupos y se creó el nombre progesterona (cetona esteroidea progestacional). [17] [176]
Uso veterinario
El uso de pruebas de progesterona en la cría de perros para determinar la ovulación se está haciendo cada vez más común. Hay varias pruebas disponibles, pero la prueba más confiable es un análisis de sangre en el que un veterinario extrae la sangre y la envía a un laboratorio para su procesamiento. Los resultados generalmente se pueden obtener en 24 a 72 horas. La razón para usar pruebas de progesterona es que el aumento de los números comienza cerca del pico preovulatorio de gonadotropinas y continúa durante la ovulación y el estro. Cuando los niveles de progesterona alcanzan ciertos niveles, pueden indicar la etapa del estro en la que se encuentra la hembra. La predicción de la fecha de nacimiento de la camada pendiente puede ser muy precisa si se conoce la fecha de ovulación. Los cachorros nacen con uno o dos días de gestación de 9 semanas en la mayoría de los casos. Sin embargo, no es posible determinar el embarazo utilizando pruebas de progesterona una vez que se ha producido un apareamiento. Esto se debe al hecho de que, en los perros, los niveles de progesterona permanecen elevados durante todo el período de estro. [177]
Precios
El precio de la progesterona puede variar según la ubicación, la cobertura del seguro, los cupones de descuento, la cantidad, la escasez, los fabricantes, las versiones de marca o genéricas, las diferentes farmacias, etc. Actualmente, 30 cápsulas de 100 mg de la versión genérica, Prometrium, de CVS Pharmacy cuestan alrededor de $40 sin ningún descuento ni seguro aplicado. La versión de marca, Progesterona, cuesta alrededor de $450 por 30 cápsulas sin ningún descuento ni seguro aplicado. [178] En comparación, Walgreens ofrece 30 cápsulas de 100 mg en la versión genérica por $51 sin seguro ni cupones aplicados. La versión de marca cuesta alrededor de $431 por 30 cápsulas de 100 mg. [179]
Referencias
Este artículo incorpora texto disponible bajo la licencia CC BY-SA 3.0.
^ ab Jameson JL, De Groot LJ (25 de febrero de 2015). Endocrinología: libro electrónico para adultos y niños. Elsevier Health Sciences. pág. 2179. ISBN 978-0-323-32195-2Archivado desde el original el 14 de enero de 2023 . Consultado el 29 de octubre de 2017 .
^ Adler N, Pfaff D, Goy RW (6 de diciembre de 2012). Handbook of Behavioral Neurobiology Volumen 7 Reproducción (1.ª ed.). Nueva York: Plenum Press. pág. 189. ISBN978-1-4684-4834-4Archivado desde el original el 14 de enero de 2023 . Consultado el 4 de julio de 2015 .
^ "progesterona (CHEBI:17026)". ChEBI . Laboratorio Europeo de Biología Molecular-EBI. Archivado desde el original el 20 de marzo de 2016 . Consultado el 4 de julio de 2015 .
^ "Progesterona_msds". Archivado desde el original el 12 de febrero de 2021 . Consultado el 19 de abril de 2018 .
^ ab Stanczyk FZ (septiembre de 2002). "Farmacocinética y potencia de las progestinas utilizadas para la terapia de reemplazo hormonal y la anticoncepción". Reseñas en Endocrine & Metabolic Disorders . 3 (3): 211–224. doi :10.1023/A:1020072325818. PMID 12215716. S2CID 27018468.
^ abc Simon JA, Robinson DE, Andrews MC, Hildebrand JR, Rocci ML, Blake RE, Hodgen GD (julio de 1993). "Absorción de progesterona micronizada oral: efecto de los alimentos, proporcionalidad de la dosis y comparación con progesterona intramuscular". Fertilidad y esterilidad . 60 (1): 26–33. doi : 10.1016/S0015-0282(16)56031-2 . PMID 8513955.
^ Fritz MA, Speroff L (28 de marzo de 2012). Endocrinología ginecológica clínica e infertilidad. Lippincott Williams & Wilkins. pp. 44–. ISBN978-1-4511-4847-3.
^ Marshall WJ, Bangert SK (2008). Química clínica. Elsevier Health Sciences. págs. 192–. ISBN978-0-7234-3455-9Archivado desde el original el 14 de enero de 2023 . Consultado el 5 de octubre de 2016 .
^ Yamazaki H, Shimada T (octubre de 1997). "Hidroxilación de progesterona y testosterona por los citocromos P450 2C19, 2C9 y 3A4 en microsomas hepáticos humanos". Archivos de bioquímica y biofísica . 346 (1): 161–169. doi :10.1006/abbi.1997.0302. PMID 9328296.
^ McKay GA, Walters MR (6 de febrero de 2013). Notas de la conferencia: Farmacología clínica y terapéutica. John Wiley & Sons. pág. 33. ISBN978-1-118-34489-7Archivado desde el original el 14 de enero de 2023 . Consultado el 27 de junio de 2015 .
^ Zutshi V, Rathore AM, Sharma K (1 de enero de 2005). Hormonas en obstetricia y ginecología. Jaypee Brothers Publishers. pág. 74. ISBN978-81-8061-427-9.[ enlace muerto permanente ]
^ ab Cometti B (noviembre de 2015). "Desarrollo farmacéutico y clínico de una nueva formulación de progesterona". Acta Obstetricia et Gynecologica Scandinavica . 94 (Supl 161): 28–37. doi : 10.1111/aogs.12765 . PMID 26342177. S2CID 31974637.
^ abc King TL, Brucker MC (25 de octubre de 2010). Farmacología para la salud de la mujer. Jones & Bartlett Publishers. págs. 372–373. ISBN978-1-4496-5800-7.
^ ab Baulieu E, Schumacher M (2000). "La progesterona como neuroesteroide neuroactivo, con especial referencia al efecto de la progesterona sobre la mielinización". Esteroides . 65 (10–11): 605–612. doi :10.1016/s0039-128x(00)00173-2. PMID 11108866. S2CID 14952168.
^ Prior JC (abril de 2019). "La progesterona es importante para la terapia de mujeres transgénero: aplicación de evidencia sobre los beneficios de la progesterona en mujeres cis". The Journal of Clinical Endocrinology and Metabolism . 104 (4): 1181–1186. doi : 10.1210/jc.2018-01777 . PMID 30608551. S2CID 58620122. Se ha acumulado evidencia de que la progesterona normal (y la ovulación), así como los niveles fisiológicos de estradiol, son necesarios durante los ciclos menstruales premenopáusicos de las mujeres cis para la fertilidad actual y la salud a largo plazo; las mujeres transgénero pueden requerir terapia con progesterona y posibles beneficios fisiológicos similares
^ Fischer J, Ganellin CR (2006). Descubrimiento de fármacos basado en análogos. John Wiley & Sons. pág. 47X. ISBN9783527607495.
^ abcdefgh Josimovich JB (11 de noviembre de 2013). Endocrinología ginecológica. Springer Science & Business Media. págs. 9, 25–29. ISBN978-1-4613-2157-6Archivado desde el original el 14 de enero de 2023 . Consultado el 1 de febrero de 2016 .
^ Thomas P, Pang Y (2012). "Receptores de progesterona de membrana: evidencia de funciones neuroprotectoras, de señalización de neuroesteroides y neuroendocrinas en células neuronales". Neuroendocrinología . 96 (2): 162–171. doi :10.1159/000339822. PMC 3489003 . PMID 22687885.
^ Valadez-Cosmes P, Vázquez-Martínez ER, Cerbón M, Camacho-Arroyo I (octubre de 2016). "Receptores de progesterona de membrana en reproducción y cáncer". Endocrinología molecular y celular . 434 : 166–175. doi :10.1016/j.mce.2016.06.027. PMID 27368976. S2CID 3826650.
^ Meyer C, Schmid R, Schmieding K, Falkenstein E, Wehling M (febrero de 1998). "Caracterización de proteínas de membrana de alta afinidad que se unen a la progesterona mediante antisuero antipéptido". Esteroides . 63 (2): 111–116. doi :10.1016/s0039-128x(97)00143-8. PMID 9516722. S2CID 40096058.
^ Kabe Y, Handa H, Suematsu M (julio de 2018). "Función y regulación estructural de la proteína de membrana sensible al monóxido de carbono (CO) PGRMC1". Journal of Clinical Biochemistry and Nutrition . 63 (1): 12–17. doi :10.3164/jcbn.17-132. PMC 6064819 . PMID 30087538.
^ Ryu CS, Klein K, Zanger UM (27 de marzo de 2017). "Receptores de progesterona asociados a la membrana: proteínas promiscuas con funciones pleiotrópicas: enfoque en las interacciones con los citocromos P450". Frontiers in Pharmacology . 8 : 159. doi : 10.3389/fphar.2017.00159 . PMC 5366339 . PMID 28396637.
^ Maurice T, Urani A, Phan VL, Romieu P (noviembre de 2001). "La interacción entre los esteroides neuroactivos y la función del receptor sigma1: consecuencias conductuales y oportunidades terapéuticas". Brain Research. Brain Research Reviews . 37 (1–3): 116–132. doi :10.1016/s0165-0173(01)00112-6. PMID 11744080. S2CID 44931783.
^ Johannessen M, Fontanilla D, Mavlyutov T, Ruoho AE, Jackson MB (febrero de 2011). "Acción antagonista de la progesterona en los receptores σ en la modulación de los canales de sodio dependientes del voltaje". Revista estadounidense de fisiología. Fisiología celular . 300 (2): C328–C337. doi :10.1152/ajpcell.00383.2010. PMC 3043630. PMID 21084640 .
^ ab Rupprecht R, Reul JM, van Steensel B, Spengler D, Söder M, Berning B, et al. (octubre de 1993). "Caracterización farmacológica y funcional de los ligandos de los receptores de glucocorticoides y mineralocorticoides humanos". Revista Europea de Farmacología . 247 (2): 145–154. doi :10.1016/0922-4106(93)90072-H. PMID 8282004.
^ Elger W, Beier S, Pollow K, Garfield R, Shi SQ, Hillisch A (noviembre de 2003). "Concepción y perfil farmacodinámico de la drospirenona". Esteroides . 68 (10–13): 891–905. doi :10.1016/j.steroids.2003.08.008. PMID 14667981. S2CID 41756726.
^ Attardi BJ, Zeleznik A, Simhan H, Chiao JP, Mattison DR, Caritis SN (diciembre de 2007). "Comparación de la unión de los receptores de progesterona y glucocorticoides y la estimulación de la expresión génica por progesterona, caproato de 17-alfa hidroxiprogesterona y progestinas relacionadas". American Journal of Obstetrics and Gynecology . 197 (6): 599.e1–599.e7. doi :10.1016/j.ajog.2007.05.024. PMC 2278032 . PMID 18060946.
^ Lei K, Chen L, Georgiou EX, Sooranna SR, Khanjani S, Brosens JJ, et al. (2012). "La progesterona actúa a través del receptor de glucocorticoides nuclear para suprimir la expresión de COX-2 inducida por IL-1β en células miometriales humanas a término". PLOS ONE . 7 (11): e50167. Bibcode :2012PLoSO...750167L. doi : 10.1371/journal.pone.0050167 . PMC 3509141 . PMID 23209664.
^ Paul SM, Purdy RH (marzo de 1992). "Esteroides neuroactivos". FASEB Journal . 6 (6): 2311–2322. doi : 10.1096/fasebj.6.6.1347506 . PMID 1347506. S2CID 221753076.
^ Kliewer SA, Goodwin B, Willson TM (octubre de 2002). "El receptor nuclear de pregnano X: un regulador clave del metabolismo xenobiótico". Endocrine Reviews . 23 (5): 687–702. doi : 10.1210/er.2001-0038 . PMID 12372848.
^ Lehmann JM, McKee DD, Watson MA, Willson TM, Moore JT, Kliewer SA (septiembre de 1998). "El receptor nuclear huérfano humano PXR es activado por compuestos que regulan la expresión del gen CYP3A4 y causan interacciones farmacológicas". The Journal of Clinical Investigation . 102 (5): 1016–1023. doi :10.1172/JCI3703. PMC 508967 . PMID 9727070.
^ Meanwell NA (8 de diciembre de 2014). Tácticas en el diseño de fármacos contemporáneos. Springer. pp. 161–. ISBN978-3-642-55041-6Archivado desde el original el 14 de enero de 2023 . Consultado el 1 de febrero de 2016 .
^ ab Legato MJ, Bilezikian JP (2004). Principios de la medicina específica de género. Gulf Professional Publishing. págs. 146–. ISBN978-0-12-440906-4Archivado desde el original el 14 de enero de 2023 . Consultado el 1 de febrero de 2016 .
^ Williams DA (24 de enero de 2012). "Metabolismo de fármacos". En Lemke TL, Williams DA (eds.). Principios de química medicinal de Foye . Lippincott Williams & Wilkins. pág. 164. ISBN978-1-60913-345-0.
^ Estrógenos: avances en investigación y aplicación: edición de 2013: ScholarlyBrief. ScholarlyEditions. 21 de junio de 2013. pp. 4–. ISBN978-1-4816-7550-5Archivado desde el original el 14 de enero de 2023 . Consultado el 1 de febrero de 2016 .
^ Strünker T, Goodwin N, Brenker C, Kashikar ND, Weyand I, Seifert R, Kaupp UB (marzo de 2011). "El canal CatSper media la entrada de Ca2+ inducida por progesterona en el esperma humano". Nature . 471 (7338): 382–386. Bibcode :2011Natur.471..382S. doi :10.1038/nature09769. PMID 21412338. S2CID 4431334.
^ Lishko PV, Botchkina IL, Kirichok Y (marzo de 2011). "La progesterona activa el canal principal de Ca2+ del esperma humano". Nature . 471 (7338): 387–391. Bibcode :2011Natur.471..387L. doi :10.1038/nature09767. PMID 21412339. S2CID 4340309.
^ Prior JC (2020). "El sistema reproductivo de la mujer como acción equilibrada del estradiol y la progesterona: un concepto revolucionario que cambia paradigmas en la salud de la mujer". Drug Discovery Today: Disease Models . 32 (Part B): 31–40. doi : 10.1016/j.ddmod.2020.11.005 .
^Kastner P, Krust A, Turcotte B, Stropp U, Tora L, Gronemeyer H, Chambon P (May 1990). "Two distinct estrogen-regulated promoters generate transcripts encoding the two functionally different human progesterone receptor forms A and B". The EMBO Journal. 9 (5): 1603–1614. doi:10.1002/j.1460-2075.1990.tb08280.x. PMC 551856. PMID 2328727.
^ a bCline JM, Wood CE (1 January 2006). Hallam SZ, Osuch JR (eds.). "Hormonal effects on the mammary gland of postmenopausal nonhuman primates". Breast Disease. 24. IOS Press: 59–70. doi:10.3233/bd-2006-24105. ISBN 978-1-58603-653-9. PMID 16917139. Archived from the original on 27 November 2023. Retrieved 2 August 2023.
^ a bJohnson LR (2003). Essential Medical Physiology. Academic Press. p. 770. ISBN 978-0-12-387584-6. Archived from the original on 14 January 2023. Retrieved 1 February 2016.
^ a bCoad J, Dunstall M (2011). Anatomy and Physiology for Midwives, with Pageburst online access,3: Anatomy and Physiology for Midwives. Elsevier Health Sciences. p. 413. ISBN 978-0-7020-3489-3.
^Landau RL, Bergenstal DM, Lugibihl K, Kascht ME (October 1955). "The metabolic effects of progesterone in man". The Journal of Clinical Endocrinology and Metabolism. 15 (10): 1194–1215. doi:10.1210/jcem-15-10-1194. PMID 13263410.
^ a b cMasiutin M, Yadav M (2023). "Alternative androgen pathways". WikiJournal of Medicine. 10: X. doi:10.15347/WJM/2023.003. S2CID 257943362.
^ a bO'Shaughnessy PJ, Antignac JP, Le Bizec B, Morvan ML, Svechnikov K, Söder O, et al. (February 2019). "Alternative (backdoor) androgen production and masculinization in the human fetus". PLOS Biology. 17 (2): e3000002. doi:10.1371/journal.pbio.3000002. PMC 6375548. PMID 30763313.
^Flück CE, Pandey AV (May 2014). "Steroidogenesis of the testis -- new genes and pathways". Annales d'Endocrinologie. 75 (2): 40–47. doi:10.1016/j.ando.2014.03.002. PMID 24793988.
^Zachmann M (February 1996). "Prismatic cases: 17,20-desmolase (17,20-lyase) deficiency". The Journal of Clinical Endocrinology and Metabolism. 81 (2): 457–459. doi:10.1210/jcem.81.2.8636249. PMID 8636249.
^Correia JN, Conner SJ, Kirkman-Brown JC (May 2007). "Non-genomic steroid actions in human spermatozoa. "Persistent tickling from a laden environment"". Seminars in Reproductive Medicine. 25 (3): 208–219. doi:10.1055/s-2007-973433. PMID 17447210. S2CID 260318879.
^Kirkman-Brown JC, Bray C, Stewart PM, Barratt CL, Publicover SJ (June 2000). "Biphasic elevation of [Ca(2+)](i) in individual human spermatozoa exposed to progesterone". Developmental Biology. 222 (2): 326–335. doi:10.1006/dbio.2000.9729. PMID 10837122.
^Kirkman-Brown JC, Barratt CL, Publicover SJ (March 2004). "Slow calcium oscillations in human spermatozoa". The Biochemical Journal. 378 (Pt 3): 827–832. doi:10.1042/BJ20031368. PMC 1223996. PMID 14606954.
^Harper CV, Barratt CL, Publicover SJ (October 2004). "Stimulation of human spermatozoa with progesterone gradients to simulate approach to the oocyte. Induction of [Ca(2+)](i) oscillations and cyclical transitions in flagellar beating". The Journal of Biological Chemistry. 279 (44): 46315–46325. doi:10.1074/jbc.M401194200. PMID 15322137.
^Marieb E (2013). Anatomy & physiology. Benjamin-Cummings. p. 903. ISBN 9780321887603.
^Tosti E, Di Cosmo A, Cuomo A, Di Cristo C, Gragnaniello G (May 2001). "Progesterone induces activation in Octopus vulgaris spermatozoa". Molecular Reproduction and Development. 59 (1): 97–105. doi:10.1002/mrd.1011. PMID 11335951. S2CID 28390608.
^ a bBowen R (6 August 2000). "Placental Hormones". Archived from the original on 17 May 2007. Retrieved 12 March 2008.
^Patel B, Elguero S, Thakore S, Dahoud W, Bedaiwy M, Mesiano S (2014). "Role of nuclear progesterone receptor isoforms in uterine pathophysiology". Human Reproduction Update. 21 (2): 155–173. doi:10.1093/humupd/dmu056. PMC 4366574. PMID 25406186.
^ a bDi Renzo GC, Giardina I, Clerici G, Brillo E, Gerli S (July 2016). "Progesterone in normal and pathological pregnancy". Hormone Molecular Biology and Clinical Investigation. 27 (1): 35–48. doi:10.1515/hmbci-2016-0038. PMID 27662646. S2CID 32239449.
^Care A, Nevitt SJ, Medley N, Donegan S, Good L, Hampson L, et al. (February 2022). "Interventions to prevent spontaneous preterm birth in women with singleton pregnancy who are at high risk: systematic review and network meta-analysis". BMJ. 376: e064547. doi:10.1136/bmj-2021-064547. PMC 8845039. PMID 35168930.
^ a b cHilton HN, Graham JD, Clarke CL (September 2015). "Minireview: Progesterone Regulation of Proliferation in the Normal Human Breast and in Breast Cancer: A Tale of Two Scenarios?". Molecular Endocrinology. 29 (9): 1230–1242. doi:10.1210/me.2015-1152. PMC 5414684. PMID 26266959.
^Barbieri RL (13 September 2013). "The Breast". In Strauss JF, Barbieri RL (eds.). Yen and Jaffe's Reproductive Endocrinology. Elsevier Health Sciences. pp. 236–. ISBN 978-1-4557-2758-2. Archived from the original on 14 January 2023. Retrieved 1 February 2016.
^Scaling AL, Prossnitz ER, Hathaway HJ (June 2014). "GPER mediates estrogen-induced signaling and proliferation in human breast epithelial cells and normal and malignant breast". Hormones & Cancer. 5 (3): 146–160. doi:10.1007/s12672-014-0174-1. PMC 4091989. PMID 24718936.
^ a b c d eAupperlee MD, Leipprandt JR, Bennett JM, Schwartz RC, Haslam SZ (May 2013). "Amphiregulin mediates progesterone-induced mammary ductal development during puberty". Breast Cancer Research. 15 (3): R44. doi:10.1186/bcr3431. PMC 3738150. PMID 23705924.
^ a bKuhl H, Schneider HP (August 2013). "Progesterone--promoter or inhibitor of breast cancer". Climacteric. 16 (Suppl 1): 54–68. doi:10.3109/13697137.2013.768806. PMID 23336704. S2CID 20808536.
^ a b cTrabert B, Sherman ME, Kannan N, Stanczyk FZ (April 2020). "Progesterone and Breast Cancer". Endocrine Reviews. 41 (2): 320–344. doi:10.1210/endrev/bnz001. PMC 7156851. PMID 31512725.
^Collaborative Group on Hormonal Factors in Breast Cancer (September 2019). "Type and timing of menopausal hormone therapy and breast cancer risk: individual participant meta-analysis of the worldwide epidemiological evidence". Lancet. 394 (10204): 1159–1168. doi:10.1016/S0140-6736(19)31709-X. PMC 6891893. PMID 31474332.
^Stute P, Wildt L, Neulen J (April 2018). "The impact of micronized progesterone on breast cancer risk: a systematic review" (PDF). Climacteric. 21 (2): 111–122. doi:10.1080/13697137.2017.1421925. PMID 29384406. S2CID 3642971. Archived (PDF) from the original on 2 February 2024. Retrieved 2 February 2024.
^Asi N, Mohammed K, Haydour Q, Gionfriddo MR, Vargas OL, Prokop LJ, et al. (July 2016). "Progesterone vs. synthetic progestins and the risk of breast cancer: a systematic review and meta-analysis". Systematic Reviews. 5 (1): 121. doi:10.1186/s13643-016-0294-5. PMC 4960754. PMID 27456847.
^Gompel A, Plu-Bureau G (August 2018). "Progesterone, progestins and the breast in menopause treatment". Climacteric. 21 (4): 326–332. doi:10.1080/13697137.2018.1476483. PMID 29852797. S2CID 46922084.
^Davey DA (October 2018). "Menopausal hormone therapy: a better and safer future". Climacteric. 21 (5): 454–461. doi:10.1080/13697137.2018.1439915. PMID 29526116. S2CID 3850275.
^ a b c d e f g hRaine-Fenning NJ, Brincat MP, Muscat-Baron Y (2003). "Skin aging and menopause : implications for treatment". American Journal of Clinical Dermatology. 4 (6): 371–378. doi:10.2165/00128071-200304060-00001. PMID 12762829. S2CID 20392538.
^ a b c d e f g hHolzer G, Riegler E, Hönigsmann H, Farokhnia S, Schmidt JB (September 2005). "Effects and side-effects of 2% progesterone cream on the skin of peri- and postmenopausal women: results from a double-blind, vehicle-controlled, randomized study". The British Journal of Dermatology. 153 (3): 626–634. doi:10.1111/j.1365-2133.2005.06685.x. PMID 16120154. S2CID 6077829.
^King SR (9 November 2012). Neurosteroids and the Nervous System. Springer Science & Business Media. pp. 44–46. ISBN 978-1-4614-5559-2.
^Fleischman DS, Fessler DM, Cholakians AE (July 2015). "Testing the Affiliation Hypothesis of Homoerotic Motivation in Humans: The Effects of Progesterone and Priming". Archives of Sexual Behavior. 44 (5): 1395–1404. doi:10.1007/s10508-014-0436-6. PMID 25420899. S2CID 9864224. Archived from the original on 23 September 2020. Retrieved 2 August 2023.
^"Homosexuality may help us bond". UoP News. Archived from the original on 2 July 2019. Retrieved 2 July 2019.
^"Having homosexual thoughts 'is an essential part of human evolution' study suggests". The Telegraph. 25 November 2014. Archived from the original on 15 February 2018. Retrieved 4 April 2018.
^"New Study Identifies Evolutionary Basis Of Homosexuality". HuffPost. 26 November 2014. Archived from the original on 27 November 2023. Retrieved 21 October 2023.
^ a b cHanukoglu I, Karavolas HJ, Goy RW (April 1977). "Progesterone metabolism in the pineal, brain stem, thalamus and corpus callosum of the female rat". Brain Research. 125 (2): 313–324. doi:10.1016/0006-8993(77)90624-2. PMID 558037. S2CID 35814845. Archived from the original on 5 March 2021. Retrieved 28 June 2019.
^Schumacher M, Guennoun R, Robert F, Carelli C, Gago N, Ghoumari A, et al. (June 2004). "Local synthesis and dual actions of progesterone in the nervous system: neuroprotection and myelination". Growth Hormone & IGF Research. 14 (Suppl A): S18–S33. doi:10.1016/j.ghir.2004.03.007. PMID 15135772.
^Singh M, Su C, Ng S (September 2013). "Non-genomic mechanisms of progesterone action in the brain". Frontiers in Neuroscience. 7: 159. doi:10.3389/fnins.2013.00159. PMC 3776940. PMID 24065876.
^Roof RL, Hall ED (May 2000). "Gender differences in acute CNS trauma and stroke: neuroprotective effects of estrogen and progesterone". Journal of Neurotrauma. 17 (5): 367–388. doi:10.1089/neu.2000.17.367. PMID 10833057.
^Pan DS, Liu WG, Yang XF, Cao F (October 2007). "Inhibitory effect of progesterone on inflammatory factors after experimental traumatic brain injury". Biomedical and Environmental Sciences. 20 (5): 432–438. PMID 18188998.
^Jiang C, Zuo F, Wang Y, Wan J, Yang Z, Lu H, et al. (June 2016). "Progesterone exerts neuroprotective effects and improves long-term neurologic outcome after intracerebral hemorrhage in middle-aged mice". Neurobiology of Aging. 42: 13–24. doi:10.1016/j.neurobiolaging.2016.02.029. PMC 4857017. PMID 27143417.
^ a bLuoma JI, Stern CM, Mermelstein PG (August 2012). "Progesterone inhibition of neuronal calcium signaling underlies aspects of progesterone-mediated neuroprotection". The Journal of Steroid Biochemistry and Molecular Biology. 131 (1–2): 30–36. doi:10.1016/j.jsbmb.2011.11.002. PMC 3303940. PMID 22101209.
^ abc Stein DG (marzo de 2008). "La progesterona ejerce efectos neuroprotectores después de una lesión cerebral". Brain Research Reviews . 57 (2): 386–397. doi :10.1016/j.brainresrev.2007.06.012. PMC 2699575 . PMID 17826842.
^ Espinoza TR, Wright DW (2011). "El papel de la progesterona en la lesión cerebral traumática". Revista de rehabilitación de traumatismos craneales . 26 (6): 497–499. doi :10.1097/HTR.0b013e31823088fa. PMC 6025750. PMID 22088981 .
^ Jiang C, Zuo F, Wang Y, Lu H, Yang Q, Wang J (enero de 2016). "La progesterona modifica la expresión de VEGF y BDNF y promueve la neurogénesis después de un accidente cerebrovascular isquémico". Neurobiología molecular . 54 (1): 571–581. doi :10.1007/s12035-015-9651-y. PMC 4938789 . PMID 26746666.
^ Herson PS, Koerner IP, Hurn PD (mayo de 2009). "Sexo, esteroides sexuales y lesión cerebral". Seminarios en Medicina Reproductiva . 27 (3): 229–239. doi :10.1055/s-0029-1216276. PMC 2675922 . PMID 19401954.
^ Li Z, Wang B, Kan Z, Zhang B, Yang Z, Chen J, et al. (enero de 2012). "La progesterona aumenta las células progenitoras endoteliales circulantes e induce la regeneración neuronal después de una lesión cerebral traumática en ratas envejecidas". Journal of Neurotrauma . 29 (2): 343–353. doi :10.1089/neu.2011.1807. PMC 3261789 . PMID 21534727.
^ ab Lynch WJ, Sofuoglu M (diciembre de 2010). "El papel de la progesterona en la adicción a la nicotina: evidencia desde el inicio hasta la recaída". Psicofarmacología experimental y clínica . 18 (6): 451–461. doi :10.1037/a0021265. PMC 3638762 . PMID 21186920.
^ Cosgrove KP, Esterlis I, McKee SA, Bois F, Seibyl JP, Mazure CM, et al. (abril de 2012). "Diferencias de sexo en la disponibilidad de receptores de acetilcolina β2*-nicotínicos en fumadores de tabaco recientemente abstinentes". Archivos de psiquiatría general . 69 (4): 418–427. doi :10.1001/archgenpsychiatry.2011.1465. PMC 3508698 . PMID 22474108.
^ Mello NK, Knudson IM, Kelly M, Fivel PA, Mendelson JH (octubre de 2011). "Efectos de la progesterona y la testosterona en la autoadministración de cocaína y la discriminación de la cocaína por parte de hembras de monos rhesus". Neuropsicofarmacología . 36 (11): 2187–2199. doi :10.1038/npp.2011.130. PMC 3176575 . PMID 21796112.
^ Buser T (1 de junio de 2012). "El impacto del ciclo menstrual y los anticonceptivos hormonales en la competitividad" (PDF) . Journal of Economic Behavior & Organization . Diferencias de género en la aversión al riesgo y la competencia. 83 (1): 1–10. doi :10.1016/j.jebo.2011.06.006. ISSN 0167-2681. Archivado (PDF) desde el original el 2 de febrero de 2024 . Consultado el 2 de febrero de 2024 .
^ Sriram D (2007). Química medicinal . Nueva Delhi: Dorling Kindersley India Pvt. Ltd. pág. 432. ISBN978-81-317-0031-0.
^ ab Blackburn S (14 de abril de 2014). Fisiología materna, fetal y neonatal. Elsevier Health Sciences. págs. 92–. ISBN978-0-323-29296-2.
^ Faivre EJ, Lange CA (enero de 2007). "Los receptores de progesterona regulan positivamente Wnt-1 para inducir la transactivación del receptor del factor de crecimiento epidérmico y la activación sostenida dependiente de c-Src de la proteína quinasa activada por mitógeno Erk1/2 en células de cáncer de mama". Biología molecular y celular . 27 (2): 466–480. doi :10.1128/MCB.01539-06. PMC 1800800 . PMID 17074804.
^ Nosek TM. "Sección 5/5ch9/s5ch9_13". Fundamentos de fisiología humana . Archivado desde el original el 24 de marzo de 2016.
^ Rothchild I (1969), Salhanick HA, Kipnis DM, Wiele RL (eds.), "La base fisiológica del efecto de aumento de temperatura de la progesterona" , Efectos metabólicos de las hormonas gonadales y los esteroides anticonceptivos , Boston, MA: Springer US, págs. 668-675, doi :10.1007/978-1-4684-1782-1_49, ISBN978-1-4684-1782-1, archivado del original el 29 de agosto de 2021 , consultado el 22 de marzo de 2021
^ Hould FS, Fried GM, Fazekas AG, Tremblay S, Mersereau WA (diciembre de 1988). "Los receptores de progesterona regulan la motilidad de la vesícula biliar". The Journal of Surgical Research . 45 (6): 505–512. doi :10.1016/0022-4804(88)90137-0. PMID 3184927.
^ "Hormonas y salud bucal". WebMD . Archivado desde el original el 18 de junio de 2016 . Consultado el 22 de julio de 2013 .
^ Picard F, Wanatabe M, Schoonjans K, Lydon J, O'Malley BW, Auwerx J (noviembre de 2002). "Los ratones knock out del receptor de progesterona tienen una homeostasis de glucosa mejorada secundaria a la proliferación de células beta". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 99 (24): 15644–15648. doi : 10.1073/pnas.202612199 . PMC 137770 . PMID 12438645.
^ Brănişteanu DD, Mathieu C (marzo de 2003). "Progesterona en diabetes mellitus gestacional: ¿culpable o no culpable?". Tendencias en endocrinología y metabolismo . 14 (2): 54–56. doi :10.1016/S1043-2760(03)00003-1. PMID 12591170. S2CID 38209977.
^ Whynott RM, Summers KM, Jakubiak M, Van Voorhis BJ, Mejia RB (junio de 2021). "El efecto del peso y el índice de masa corporal en los valores de progesterona sérica y la tasa de nacidos vivos en ciclos de fertilización in vitro criopreservados". F&S Reports . 2 (2): 195–200. doi :10.1016/j.xfre.2021.02.005. PMC 8267385 . PMID 34278354.
^ Kaemmle LM, Stadler A, Janka H, von Wolff M, Stute P (agosto de 2022). "El impacto de la progesterona micronizada en los eventos cardiovasculares: una revisión sistemática". Climaterio . 25 (4): 327–336. doi :10.1080/13697137.2021.2022644. PMID 35112635. S2CID 246487187.
^ Gasser S, Heidemeyer K, von Wolff M, Stute P (junio de 2021). "Impacto de la progesterona en la piel y el cabello en la menopausia: una revisión exhaustiva". Climaterio . 24 (3): 229–235. doi :10.1080/13697137.2020.1838476. PMID 33527841. S2CID 231757325.
^ Javed AA, Mayhew AJ, Shea AK, Raina P (agosto de 2019). "Asociación entre la terapia hormonal y la masa muscular en mujeres posmenopáusicas: una revisión sistemática y un metanálisis". JAMA Network Open . 2 (8): e1910154. doi :10.1001/jamanetworkopen.2019.10154. PMC 6716293 . PMID 31461147.
^ Coquoz A, Gruetter C, Stute P (abril de 2019). "Impacto de la progesterona micronizada en el peso corporal, el índice de masa corporal y el metabolismo de la glucosa: una revisión sistemática". Climaterio . 22 (2): 148–161. doi :10.1080/13697137.2018.1514003. PMID 30477366. S2CID 53782622.
^ Häggström M, Richfield D (2014). "Diagrama de las vías de la esteroidogénesis humana". WikiJournal of Medicine . 1 (1). doi : 10.15347/wjm/2014.005 . ISSN 2002-4436.
^ Bewick PM (2002). Productos naturales medicinales: un enfoque biosintético . Nueva York: Wiley. p. 244. ISBN0-471-49641-3.
^ Duport C, Spagnoli R, Degryse E, Pompon D (febrero de 1998). "Biosíntesis autosuficiente de pregnenolona y progesterona en levaduras modificadas genéticamente". Nature Biotechnology . 16 (2): 186–189. doi :10.1038/nbt0298-186. PMID 9487528. S2CID 852617.
^ Zavod RM (24 de enero de 2012). "Salud de la mujer". En Lemke TL, Williams DA (eds.). Principios de química medicinal de Foye . Lippincott Williams & Wilkins. págs. 1397–. ISBN978-1-60913-345-0Archivado desde el original el 14 de enero de 2023 . Consultado el 19 de julio de 2018 .
^ Progesterona - Drugs.com, archivado del original el 27 de marzo de 2019 , consultado el 23 de agosto de 2015
^ abc Falcone T, Hurd WW (2007). Medicina y cirugía reproductiva clínica. Elsevier Health Sciences. pp. 22–. ISBN978-0-323-03309-1Archivado desde el original el 10 de enero de 2023 . Consultado el 6 de noviembre de 2016 .
^ abcd Cupps PT (20 de febrero de 1991). Reproducción en animales domésticos. Elsevier. pp. 101–. ISBN978-0-08-057109-6.
^ abcdefghi Stanczyk FZ (noviembre de 2003). "No todas las progestinas son iguales". Esteroides . 68 (10–13): 879–890. doi :10.1016/j.steroids.2003.08.003. PMID 14667980. S2CID 44601264.
^ Dowd FJ, Johnson B, Mariotti A (3 de septiembre de 2016). Farmacología y terapéutica para odontología. Elsevier Health Sciences. pp. 448–. ISBN978-0-323-44595-5.
^ abcd Kuhl H (agosto de 2005). "Farmacología de los estrógenos y progestágenos: influencia de diferentes vías de administración". Climaterio . 8 (Supl 1): 3–63. doi :10.1080/13697130500148875. PMID 16112947. S2CID 24616324.
^ ab Plant TM, Zeleznik AJ (15 de noviembre de 2014). Knobil y Neill's Physiology of Reproduction. Academic Press. pp. 304–. ISBN978-0-12-397769-4.
^ ab Santoro NF, Neal-Perry G (11 de septiembre de 2010). Amenorrea: una guía clínica basada en casos. Springer Science & Business Media. pp. 13–. ISBN978-1-60327-864-5Archivado desde el original el 14 de enero de 2023 . Consultado el 6 de noviembre de 2016 .
^ Reddy DS (2010). "Neurosteroides". Diferencias sexuales en el cerebro humano, sus fundamentos e implicaciones . Progreso en la investigación cerebral. Vol. 186. Elsevier. págs. 113–37. doi :10.1016/B978-0-444-53630-3.00008-7. ISBN9780444536303. PMC 3139029 . PMID 21094889.
^ ab Baulieu E, Kelly PA (30 de noviembre de 1990). Hormonas: de las moléculas a la enfermedad. Springer Science & Business Media. pp. 401–. ISBN978-0-412-02791-8.
^ Beranič N, Gobec S, Rižner TL (mayo de 2011). "Progestinas como inhibidores de las 20-cetosteroides reductasas humanas, AKR1C1 y AKR1C3". Interacciones químico-biológicas . 191 (1–3): 227–233. Bibcode :2011CBI...191..227B. doi :10.1016/j.cbi.2010.12.012. PMID 21182831.
^ Anderson GD, Odegard PS (octubre de 2004). "Farmacocinética del estrógeno y la progesterona en la enfermedad renal crónica". Avances en la enfermedad renal crónica . 11 (4): 357–360. doi :10.1053/j.ackd.2004.07.001. PMID 15492972.
^ Greenblatt JM, Brogan K (27 de abril de 2016). Terapias integradoras para la depresión: redefinición de modelos de evaluación, tratamiento y prevención. CRC Press. pp. 201–. ISBN978-1-4987-0230-0.
^ Graham C (2 de diciembre de 2012). Biología reproductiva de los grandes simios: perspectivas comparativas y biomédicas. Elsevier. pp. 179–. ISBN978-0-323-14971-6Archivado desde el original el 14 de enero de 2023 . Consultado el 6 de noviembre de 2016 .
^Strushkevich N, Gilep AA, Shen L, Arrowsmith CH, Edwards AM, Usanov SA, Park HW (February 2013). "Structural insights into aldosterone synthase substrate specificity and targeted inhibition". Molecular Endocrinology. 27 (2): 315–324. doi:10.1210/me.2012-1287. PMC 5417327. PMID 23322723.
^van Rooyen D, Gent R, Barnard L, Swart AC (April 2018). "The in vitro metabolism of 11β-hydroxyprogesterone and 11-ketoprogesterone to 11-ketodihydrotestosterone in the backdoor pathway". The Journal of Steroid Biochemistry and Molecular Biology. 178: 203–212. doi:10.1016/j.jsbmb.2017.12.014. PMID 29277707. S2CID 3700135.
^ a bde Azevedo Piccinato C (2008). Regulation of Steroid Metabolism and the Hepatic Transcriptome by Estradiol and Progesterone. pp. 24–25. ISBN 978-1-109-04632-8.[permanent dead link]
^Akalin S (January 1991). "Effects of ketoconazole in hirsute women". Acta Endocrinologica. 124 (1): 19–22. doi:10.1530/acta.0.1240019. PMID 1825737. S2CID 9831739.
^ a bAufrère MB, Benson H (June 1976). "Progesterone: an overview and recent advances". Journal of Pharmaceutical Sciences. 65 (6): 783–800. doi:10.1002/jps.2600650602. PMID 945344.
^Stricker R, Eberhart R, Chevailler MC, Quinn FA, Bischof P, Stricker R (2006). "Establishment of detailed reference values for luteinizing hormone, follicle stimulating hormone, estradiol, and progesterone during different phases of the menstrual cycle on the Abbott ARCHITECT analyzer". Clinical Chemistry and Laboratory Medicine. 44 (7): 883–887. doi:10.1515/CCLM.2006.160. PMID 16776638. S2CID 524952.
^Csapo AI, Pulkkinen MO, Wiest WG (March 1973). "Effects of luteectomy and progesterone replacement therapy in early pregnant patients". American Journal of Obstetrics and Gynecology. 115 (6): 759–765. doi:10.1016/0002-9378(73)90517-6. PMID 4688578.
^NIH Clinical Center (16 August 2004). "Progesterone Historical Reference Ranges". United States National Institutes of Health. Archived from the original on 9 January 2009. Retrieved 12 March 2008.
^Chernecky CC, Berger BJ (31 October 2012). Laboratory Tests and Diagnostic Procedures - E-Book. Elsevier Health Sciences. pp. 908–. ISBN 978-1-4557-4502-9. Archived from the original on 27 February 2024. Retrieved 23 August 2023.
^Becker KL (2001). Principles and Practice of Endocrinology and Metabolism. Lippincott Williams & Wilkins. pp. 889, 940. ISBN 978-0-7817-1750-2. Archived from the original on 27 February 2024. Retrieved 23 August 2023.
^Josimovich JB (11 November 2013). Gynecologic Endocrinology. Springer Science & Business Media. pp. 9, 25–29, 139. ISBN 978-1-4613-2157-6. Archived from the original on 14 January 2023. Retrieved 1 February 2016.
^van Keep P, Utian W (6 December 2012). The Premenstrual Syndrome: Proceedings of a workshop held during the Sixth International Congress of Psychosomatic Obstetrics and Gynecology, Berlin, September 1980. Springer Science & Business Media. pp. 51–52. ISBN 978-94-011-6255-5. Archived from the original on 14 January 2023. Retrieved 1 February 2016.
^Strauss JF, Barbieri RL (2009). Yen and Jaffe's Reproductive Endocrinology: Physiology, Pathophysiology, and Clinical Management. Elsevier Health Sciences. pp. 807–. ISBN 978-1-4160-4907-4. Archived from the original on 10 January 2023. Retrieved 23 August 2023.
^Bajaj L, Berman S (1 January 2011). Berman's Pediatric Decision Making. Elsevier Health Sciences. pp. 160–. ISBN 978-0-323-05405-8. Archived from the original on 11 January 2023. Retrieved 23 August 2023.
^Lauritzen C (1988). "Natürliche und Synthetische Sexualhormone – Biologische Grundlagen und Behandlungsprinzipien" [Natural and Synthetic Sexual Hormones – Biological Basis and Medical Treatment Principles]. In Hermann P. G. Schneider, Christian Lauritzen, Eberhard Nieschlag (eds.). Grundlagen und Klinik der Menschlichen Fortpflanzung [Foundations and Clinic of Human Reproduction] (in German). Walter de Gruyter. pp. 229–306. ISBN 978-3110109689. OCLC 35483492. Archived from the original on 1 October 2023. Retrieved 23 August 2023.
^Little, A. B., & Billiar, R. B. (1983). Progestagens. In Endocrinology of Pregnancy, 3rd Edition (pp. 92–111). Harper and Row Philadelphia. https://scholar.google.com/scholar?cluster=2512291948467467634 Archived 22 February 2022 at the Wayback Machine
^ a b c d e f g h iProgesterone Reference Ranges, Performed at the Clinical Center at the National Institutes of Health, Bethesda MD, 03Feb09
^ a b c d e f g hConverted from mass values using molar mass of 314.46 g/mol
^Häggström M (2014). "Reference ranges for estradiol, progesterone, luteinizing hormone and follicle-stimulating hormone during the menstrual cycle". WikiJournal of Medicine. 1 (1). doi:10.15347/wjm/2014.001. ISSN 2002-4436. S2CID 88035135.
^Goodson III WH, Handagama P, Moore II DH, Dairkee S (13 December 2007). "Milk products are a source of dietary progesterone". 30th Annual San Antonio Breast Cancer Symposium. pp. abstract # 2028. Archived from the original on 20 September 2008. Retrieved 12 March 2008.
^Pauli GF, Friesen JB, Gödecke T, Farnsworth NR, Glodny B (March 2010). "Occurrence of progesterone and related animal steroids in two higher plants". Journal of Natural Products. 73 (3): 338–345. doi:10.1021/np9007415. PMID 20108949. S2CID 26467578.
^Applezweig N (May 1969). "Steroids". Chemical Week. 104: 57–72. PMID 12255132.
^Noguchi E, Fujiwara Y, Matsushita S, Ikeda T, Ono M, Nohara T (September 2006). "Metabolism of tomato steroidal glycosides in humans". Chemical & Pharmaceutical Bulletin. 54 (9): 1312–1314. doi:10.1248/cpb.54.1312. PMID 16946542.
^Yang DJ, Lu TJ, Hwang LS (October 2003). "Isolation and identification of steroidal saponins in Taiwanese yam cultivar (Dioscorea pseudojaponica Yamamoto)" (PDF). Journal of Agricultural and Food Chemistry. 51 (22): 6438–6444. doi:10.1021/jf030390j. PMID 14558759. Archived (PDF) from the original on 3 August 2022. Retrieved 2 April 2022.
^"Final report of the amended safety assessment of Dioscorea Villosa (Wild Yam) root extract". International Journal of Toxicology. 23 (Suppl 2): 49–54. 2004. doi:10.1080/10915810490499055. PMID 15513824. S2CID 962216.
^Niño J, Jiménez DA, Mosquera OM, Correa YM (2007). "Diosgenin quantification by HPLC in a Dioscorea polygonoides tuber collection from colombian flora". Journal of the Brazilian Chemical Society. 18 (5): 1073–1076. doi:10.1590/S0103-50532007000500030. S2CID 95193700.
^Myoda T, Nagai T, Nagashima T (2005). "Properties of starches in yam (Dioscorea spp.) tuber". Current Topics in Food Science and Technology: 105–114. ISBN 81-308-0003-9.
^Wesp LM, Deutsch MB (March 2017). "Hormonal and Surgical Treatment Options for Transgender Women and Transfeminine Spectrum Persons". The Psychiatric Clinics of North America. 40 (1): 99–111. doi:10.1016/j.psc.2016.10.006. PMID 28159148.
^Dolitsky SN, Cordeiro Mitchell CN, Stadler SS, Segars JH (November 2020). "Efficacy of progestin-only treatment for the management of menopausal symptoms: a systematic review". Menopause. 28 (2): 217–224. doi:10.1097/GME.0000000000001676. PMID 33109992. S2CID 225100434.
^Ruan X, Mueck AO (November 2014). "Systemic progesterone therapy--oral, vaginal, injections and even transdermal?". Maturitas. 79 (3): 248–255. doi:10.1016/j.maturitas.2014.07.009. PMID 25113944.
^Filicori M (November 2015). "Clinical roles and applications of progesterone in reproductive medicine: an overview". Acta Obstetricia et Gynecologica Scandinavica. 94 (Suppl 161): 3–7. doi:10.1111/aogs.12791. PMID 26443945.
^Ciampaglia W, Cognigni GE (November 2015). "Clinical use of progesterone in infertility and assisted reproduction". Acta Obstetricia et Gynecologica Scandinavica. 94 (Suppl 161): 17–27. doi:10.1111/aogs.12770. PMID 26345161. S2CID 40753277.
^Choi SJ (September 2017). "Use of progesterone supplement therapy for prevention of preterm birth: review of literatures". Obstetrics & Gynecology Science. 60 (5): 405–420. doi:10.5468/ogs.2017.60.5.405. PMC 5621069. PMID 28989916.
^Coomarasamy A, Harb HM, Devall AJ, Cheed V, Roberts TE, Goranitis I, et al. (June 2020). "Progesterone to prevent miscarriage in those with early pregnancy bleeding: the PRISM RCT". Health Technology Assessment. 24 (33): 1–70. doi:10.3310/hta24330. PMC 7355406. PMID 32609084.
^ a bElks J (14 November 2014). The Dictionary of Drugs: Chemical Data: Chemical Data, Structures and Bibliographies. Springer. pp. 1024–. ISBN 978-1-4757-2085-3.
^ a bIndex Nominum 2000: International Drug Directory. Taylor & Francis. January 2000. pp. 880–. ISBN 978-3-88763-075-1.
^Numazawa M, Nagaoka M, Kunitama Y (September 1986). "Regiospecific deoxygenation of the dihydroxyacetone moiety at C-17 of corticoid steroids with iodotrimethylsilane". Chemical & Pharmaceutical Bulletin. 34 (9): 3722–3726. doi:10.1248/cpb.34.3722. PMID 3815593.
^ a bMarker RE, Krueger J (1940). "Sterols. CXII. Sapogenins. XLI. The Preparation of Trillin and its Conversion to Progesterone". J. Am. Chem. Soc. 62 (12): 3349–3350. doi:10.1021/ja01869a023.
^Goswami A, Kotoky R, Rastogi RC, Ghosh AC (1 May 2003). "A One-Pot Efficient Process for 16-Dehydropregnenolone Acetate". Organic Process Research & Development. 7 (3): 306–308. doi:10.1021/op0200625.
^Heyl FW (1950). "Progesterone from 3-Acetoxybisnor-5-cholenaldehyde and 3-Ketobisnor-4-cholenaldehyde". Journal of the American Chemical Society. 72 (6): 2617–2619. doi:10.1021/ja01162a076.
^Slomp G (1958). "Ozonolysis. II. 1 The Effect of Pyridine on the Ozonolysis of 4,22-Stigmastadien-3-one 2". Journal of the American Chemical Society. 80 (4): 915–921. doi:10.1021/ja01537a041.
^Sundararaman P, Djerassi C (October 1977). "A convenient synthesis of progesterone from stigmasterol". The Journal of Organic Chemistry. 42 (22): 3633–3634. doi:10.1021/jo00442a044. PMID 915584.
^"Nova Transcripts: Forgotten Genius". PBS.org. 6 February 2007. Archived from the original on 11 October 2018. Retrieved 8 September 2017.
^"Giants of the Past". lipidlibrary.aocs.org. Archived from the original on 15 April 2012.
^ a b cJohnson WS, Gravestock MB, McCarry BE (August 1971). "Acetylenic bond participation in biogenetic-like olefinic cyclizations. II. Synthesis of dl-progesterone". Journal of the American Chemical Society. 93 (17): 4332–4334. doi:10.1021/ja00746a062. PMID 5131151.
^Corner GW, Allen WM (1 March 1929). "Physiology of the corpus luteum". American Journal of Physiology. Legacy Content. 88 (2): 326–339. doi:10.1152/ajplegacy.1929.88.2.326. ISSN 0002-9513. Archived from the original on 12 August 2021. Retrieved 12 August 2021.
^ a b cCoutinho EM, Segal SJ (1999). Is Menstruation Obsolete?. Oxford University Press. pp. 31–. ISBN 978-0-19-513021-8. Archived from the original on 14 January 2023. Retrieved 5 October 2016.
^Walker A (7 March 2008). The Menstrual Cycle. Routledge. pp. 49–. ISBN 978-1-134-71411-7.
^Piosik R (2003). "Adolf Butenandt und sein Wirken an der Technischen Hochschule Danzig". CHEMKON. 10 (3): 135–138. doi:10.1002/ckon.200390038.
^Ginsburg B (6 December 2012). Premenstrual Syndrome: Ethical and Legal Implications in a Biomedical Perspective. Springer Science & Business Media. pp. 274–. ISBN 978-1-4684-5275-4. Archived from the original on 14 January 2023. Retrieved 5 October 2016.
^Rolleston HD (1936). The Endocrine Organs in Health and Disease: With an Historical Review. Oxford University Press, H. Milford. p. 406. Archived from the original on 14 January 2023. Retrieved 5 October 2016.
^Allen WM (October 1970). "Progesterone: how did the name originate?". Southern Medical Journal. 63 (10): 1151–1155. doi:10.1097/00007611-197010000-00012. PMID 4922128. S2CID 35867375.
^Refsal K (February 2009). "Interpretation of Serum Progesterone Results for Management of Breeding in Dogs" (PDF). Webcd.endo.ref. Archived from the original on 29 August 2021. Retrieved 26 February 2018.
^"Progesterone Prices, Coupons & Savings Tips - GoodRx". www.goodrx.com. Archived from the original on 30 March 2023. Retrieved 1 August 2023.
^"Progesterone Prices, Coupons & Savings Tips - GoodRx". www.goodrx.com. Archived from the original on 30 March 2023. Retrieved 1 August 2023.