Los bastones son células fotorreceptoras de la retina del ojo que pueden funcionar mejor con poca luz que el otro tipo de fotorreceptor visual, los conos . Los bastones suelen encontrarse concentrados en los bordes externos de la retina y se utilizan en la visión periférica . En promedio, hay aproximadamente 92 millones de bastones (frente a ~6 millones de conos) en la retina humana. [1] Los bastones son más sensibles que los conos y son casi completamente responsables de la visión nocturna . Sin embargo, los bastones tienen un papel pequeño en la visión del color , que es la razón principal por la que los colores son mucho menos evidentes en la luz tenue.
Los bastones son un poco más largos y delgados que los conos, pero tienen la misma estructura básica. Los discos que contienen opsina se encuentran en el extremo de la célula adyacente al epitelio pigmentario de la retina , que a su vez está unido al interior del ojo . La estructura de disco apilado de la porción detectora de la célula permite una eficiencia muy alta. Los bastones son mucho más comunes que los conos, con alrededor de 120 millones de células de bastón en comparación con 6 a 7 millones de células de cono. [2]
Al igual que los conos, las células bastón tienen una terminal sináptica, un segmento interno y un segmento externo. La terminal sináptica forma una sinapsis con otra neurona, generalmente una célula bipolar o una célula horizontal . Los segmentos interno y externo están conectados por un cilio , [3] que recubre el segmento distal. [4] El segmento interno contiene orgánulos y el núcleo de la célula , mientras que el segmento externo del bastón (abreviado como ROS), que apunta hacia la parte posterior del ojo, contiene los materiales que absorben la luz. [3]
Una célula humana tipo bastón tiene aproximadamente 2 micrones de diámetro y 100 micrones de largo. [5] Los bastones no son todos morfológicamente iguales; en ratones, los bastones cercanos a la capa sináptica plexiforme externa muestran una longitud reducida debido a una terminal sináptica acortada. [6]

En los vertebrados, la activación de una célula fotorreceptora es una hiperpolarización (inhibición) de la célula. Cuando no están siendo estimuladas, como en la oscuridad, las células bastón y cono se despolarizan y liberan un neurotransmisor espontáneamente. Este neurotransmisor hiperpolariza la célula bipolar . Las células bipolares existen entre los fotorreceptores y las células ganglionares y actúan para transmitir señales desde los fotorreceptores a las células ganglionares . Como resultado de la hiperpolarización de la célula bipolar, no libera su transmisor en la sinapsis bipolar-ganglionar y la sinapsis no se excita.
La activación de los fotopigmentos por la luz envía una señal hiperpolarizando la célula bastón, lo que lleva a que esta no envíe su neurotransmisor, lo que lleva a que la célula bipolar libere su transmisor en la sinapsis bipolar-ganglionar y excite la sinapsis.
La despolarización de las células bastón (que provoca la liberación de su neurotransmisor) se produce porque en la oscuridad, las células tienen una concentración relativamente alta de guanosina cíclica 3'-5' monofosfato (cGMP), que abre canales iónicos (principalmente canales de sodio, aunque el calcio también puede entrar a través de estos canales). Las cargas positivas de los iones que entran en la célula a través de su gradiente electroquímico cambian el potencial de membrana de la célula , causan despolarización y conducen a la liberación del neurotransmisor glutamato . El glutamato puede despolarizar algunas neuronas e hiperpolarizar otras, lo que permite que los fotorreceptores interactúen de manera antagónica.
Cuando la luz incide sobre los pigmentos fotorreceptores de la célula fotorreceptora, el pigmento cambia de forma. El pigmento, llamado rodopsina (la conopsina se encuentra en las células de los conos), está formado por una proteína grande llamada opsina (situada en la membrana plasmática), a la que se une un grupo prostético unido covalentemente: una molécula orgánica llamada retinal (un derivado de la vitamina A ). El retinal existe en la forma 11-cis-retinal cuando está en la oscuridad, y la estimulación por la luz hace que su estructura cambie a todo-trans-retinal. Este cambio estructural provoca una mayor afinidad por la proteína reguladora llamada transducina (un tipo de proteína G). Al unirse a la rodopsina, la subunidad alfa de la proteína G reemplaza una molécula de GDP por una molécula de GTP y se activa. Este reemplazo hace que la subunidad alfa de la proteína G se disocie de las subunidades beta y gamma de la proteína G. Como resultado, la subunidad alfa ahora es libre de unirse a la fosfodiesterasa cGMP (una proteína efectora). [8] La subunidad alfa interactúa con las subunidades inhibidoras gamma de la PDE y evita que bloqueen los sitios catalíticos en las subunidades alfa y beta de la PDE, lo que lleva a la activación de la fosfodiesterasa de cGMP, que hidroliza el cGMP (el segundo mensajero), descomponiéndolo en 5'-GMP. [9] La reducción de cGMP permite que los canales iónicos se cierren, lo que evita la entrada de iones positivos, hiperpolariza la célula y detiene la liberación del neurotransmisor glutamato. [3] Aunque las células de los conos utilizan principalmente la sustancia neurotransmisora acetilcolina , las células de los bastones utilizan una variedad. Todo el proceso por el cual la luz inicia una respuesta sensorial se llama fototransducción visual.
La activación de una sola unidad de rodopsina , el pigmento fotosensible de los bastones, puede provocar una gran reacción en la célula porque la señal se amplifica. Una vez activada, la rodopsina puede activar cientos de moléculas de transducina, cada una de las cuales a su vez activa una molécula de fosfodiesterasa, que puede descomponer más de mil moléculas de cGMP por segundo. [3] Por lo tanto, los bastones pueden tener una gran respuesta a una pequeña cantidad de luz.
Como el componente retiniano de la rodopsina se deriva de la vitamina A , una deficiencia de vitamina A provoca un déficit del pigmento que necesitan los bastones. En consecuencia, menos bastones son capaces de responder de forma adecuada en condiciones de oscuridad y, como los conos están mal adaptados para ver en la oscuridad, puede producirse ceguera nocturna .
Los bastones utilizan tres mecanismos inhibidores (mecanismos de retroalimentación negativa) para permitir un rápido retorno al estado de reposo después de un destello de luz.
En primer lugar, existe una quinasa de rodopsina (RK) que fosforilaría la cola citosólica de la rodopsina activada en las múltiples serinas, inhibiendo parcialmente la activación de la transducina . Además, una proteína inhibidora, la arrestina , se une a las rodopsinas fosforiladas para inhibir aún más la actividad de la rodopsina.
Mientras que la arrestina desactiva la rodopsina, una proteína RGS (que funciona como una proteína activadora de GTPasa (GAP)) lleva a la transducina (proteína G) a un estado "apagado" al aumentar la tasa de hidrólisis del GTP unido al GDP.
Cuando la concentración de cGMP disminuye, los canales sensibles a cGMP que estaban abiertos anteriormente se cierran, lo que lleva a una reducción en la entrada de iones de calcio. La disminución asociada en la concentración de iones de calcio estimula las proteínas sensibles a los iones de calcio, que luego activan la guanilil ciclasa para reponer el cGMP, restableciéndolo rápidamente a su concentración original. Esto abre los canales sensibles a cGMP y causa una despolarización de la membrana plasmática. [10]
Cuando los bastones se exponen a una alta concentración de fotones durante un período prolongado, se desensibilizan (se adaptan) al entorno.
Como la rodopsina es fosforilada por la quinasa de la rodopsina (un miembro de las quinasas GPCR (GRK)), se une con alta afinidad a la arrestina . La arrestina unida puede contribuir al proceso de desensibilización de al menos dos maneras. En primer lugar, evita la interacción entre la proteína G y el receptor activado. En segundo lugar, sirve como proteína adaptadora para ayudar al receptor a la maquinaria de endocitosis dependiente de clatrina (para inducir la endocitosis mediada por el receptor). [10]
Una célula bastón es lo suficientemente sensible como para responder a un solo fotón de luz [11] y es aproximadamente 100 veces más sensible a un solo fotón que los conos. Dado que los bastones requieren menos luz para funcionar que los conos, son la fuente principal de información visual durante la noche ( visión escotópica ). Las células cono, por otro lado, requieren de decenas a cientos de fotones para activarse. Además, múltiples células bastón convergen en una sola interneurona , recolectando y amplificando las señales. Sin embargo, esta convergencia tiene un costo para la agudeza visual (o resolución de imagen ) porque la información agrupada de múltiples células es menos clara de lo que sería si el sistema visual recibiera información de cada célula bastón individualmente.
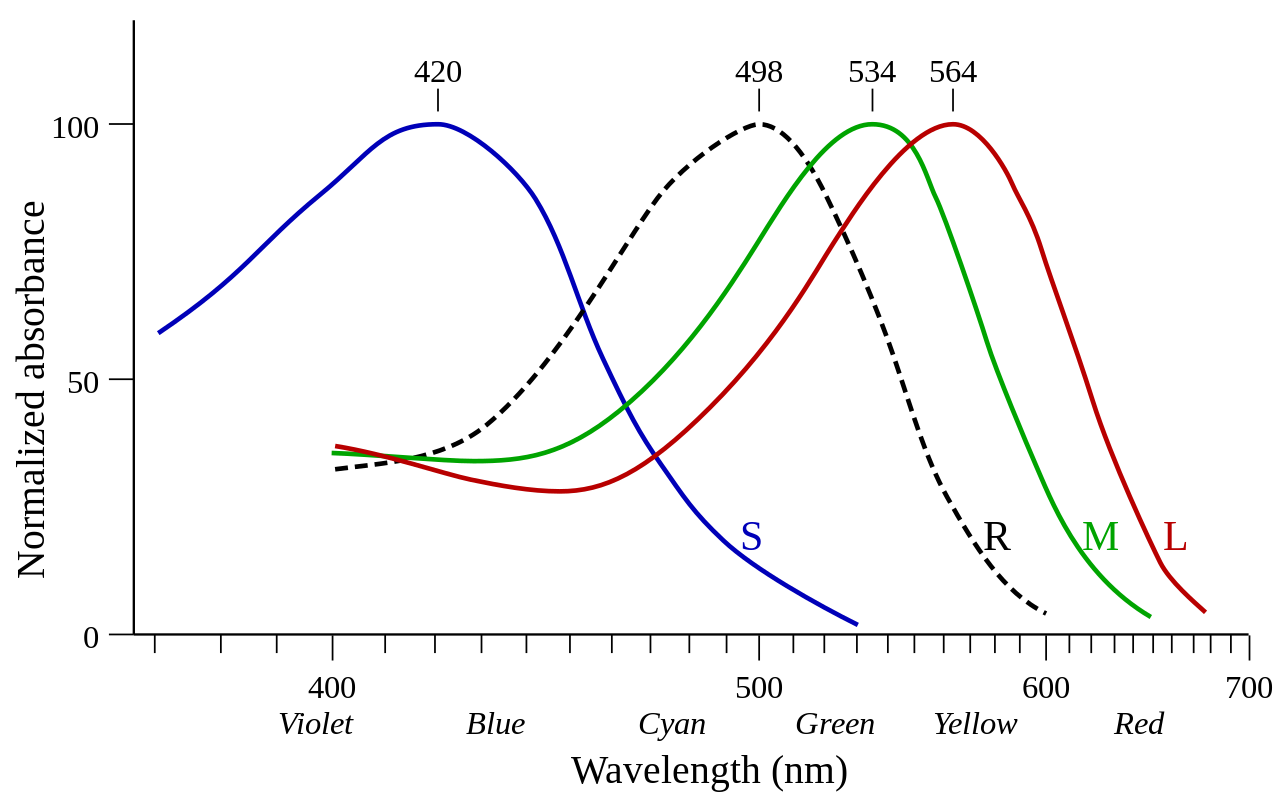
Los bastones también responden más lentamente a la luz que los conos y los estímulos que reciben se suman en un lapso de aproximadamente 100 milisegundos. Si bien esto hace que los bastones sean más sensibles a cantidades más pequeñas de luz, también significa que su capacidad para detectar cambios temporales, como imágenes que cambian rápidamente, es menos precisa que la de los conos. [3]
Los experimentos de George Wald y otros demostraron que los bastones son más sensibles a longitudes de onda de luz de alrededor de 498 nm (verde-azul) e insensibles a longitudes de onda mayores de aproximadamente 640 nm (rojo). Esto es responsable del efecto Purkinje : a medida que la intensidad disminuye en el crepúsculo, los bastones toman el control y, antes de que el color desaparezca por completo, la sensibilidad máxima de la visión se desplaza hacia la sensibilidad máxima de los bastones (azul-verde). [13]
Lista de los distintos tipos de células del cuerpo humano adulto