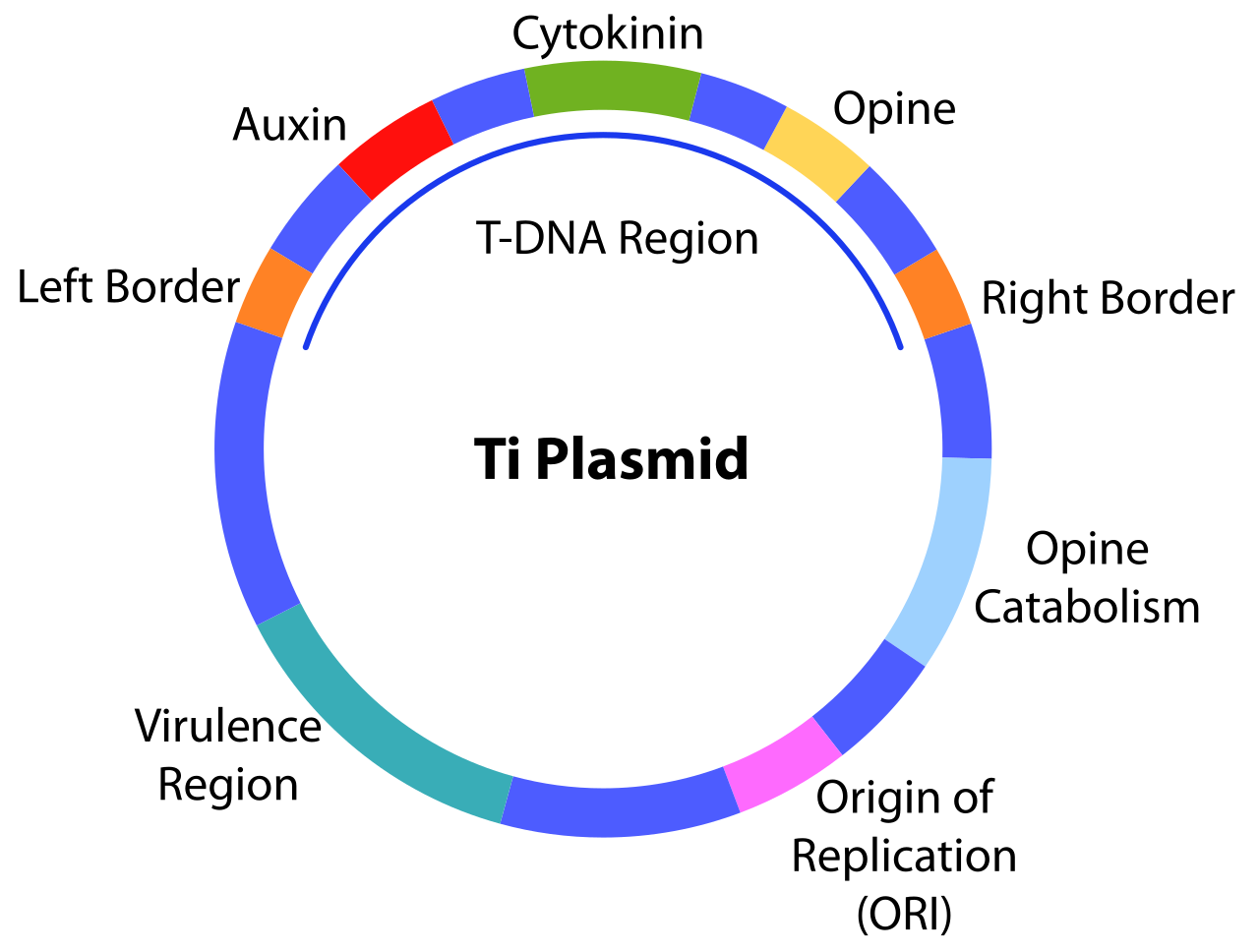
El ADN de transferencia (abreviado como ADN-T ) es el ADN transferido del plásmido inductor de tumores (Ti) de algunas especies de bacterias como Agrobacterium tumefaciens y Agrobacterium rhizogenes (en realidad, un plásmido Ri) . El ADN-T se transfiere desde la bacteria al genoma de ADN nuclear de la planta huésped . [1] La capacidad de este plásmido especializado inductor de tumores (Ti) se atribuye a dos regiones esenciales necesarias para la transferencia de ADN a la célula huésped. El ADN-T está bordeado por repeticiones de 25 pares de bases en cada extremo. La transferencia se inicia en el borde derecho y termina en el borde izquierdo y requiere los genes vir del plásmido Ti.
El ADN-T bacteriano tiene una longitud de aproximadamente 24 000 pares de bases [2] [3] y contiene genes expresados en plantas que codifican enzimas que sintetizan opinas y fitohormonas . Al transferir el ADN-T al genoma de la planta, la bacteria esencialmente reprograma las células vegetales para que crezcan hasta convertirse en un tumor y produzcan una fuente de alimento única para la bacteria. La síntesis de las hormonas vegetales auxina y citoquinina por enzimas codificadas en el ADN-T permite que la célula vegetal crezca en exceso, formando así los tumores de agalla de la corona típicamente inducidos por la infección por Agrobacterium tumefaciens . [4] Agrobacterium rhizogenes causa una infección similar conocida como enfermedad de la raíz pilosa . Las opinas son derivados de aminoácidos utilizados por la bacteria como fuente de carbono y energía. Este proceso natural de transferencia horizontal de genes en plantas se está utilizando como una herramienta para la investigación fundamental y aplicada en biología vegetal a través de la transformación de genes extraños mediada por Agrobacterium tumefaciens y la mutagénesis insercional. [5] [6] Los genomas de las plantas se pueden diseñar mediante el uso de Agrobacterium para la administración de secuencias alojadas en vectores binarios de ADN-T .
El proceso de infección del ADN-T en la célula huésped y su integración en su núcleo implica múltiples pasos. Primero, las bacterias se multiplican en la savia de la herida antes de la infección y luego se adhieren a las paredes celulares de la planta. La expresión de los genes de virulencia bacteriana de aproximadamente 10 operones se activa por la percepción de compuestos fenólicos como la acetosiringona emitida por el tejido de la planta herida y sigue al contacto célula-célula. Luego, este proceso continúa con la translocación macromolecular de Agrobacterium al citoplasma de la célula huésped, la transmisión del ADN-T junto con las proteínas asociadas (llamado complejo T ) al núcleo de la célula huésped seguido del desmontaje del complejo T, la integración estable del ADN-T en el genoma de la planta huésped y la expresión final de los genes transferidos . La integración del ADN-T en un genoma huésped implica la formación de una muesca monocatenaria en el ADN en el borde derecho del plásmido Ti. Esta muesca crea una región de ADN monocatenario desde el borde izquierdo del gen del ADN-T hasta el borde derecho que fue cortado. Luego, las proteínas de unión monocatenarias se unen al ADN monocatenario. La síntesis de ADN desplaza la región monocatenaria y luego una segunda muesca en la región del borde izquierdo libera el fragmento de ADN-T monocatenario. Además, este fragmento puede incorporarse al genoma del huésped. [7]
Se sabe que Agrobacterium ha desarrollado un sistema de control que utiliza factores hospedantes vegetales y procesos celulares para varias vías de respuesta de defensa de la planta hospedante para invadir el núcleo de la célula hospedante. Para la integración del T-ADN en el genoma hospedante objetivo, Agrobacterium lleva a cabo múltiples interacciones con factores de la planta hospedante. [7] Para interactuar con las proteínas de la planta hospedante, muchas proteínas de virulencia de Agrobacterium están codificadas por genes vir. La expresión del gen vir de Agrobacterium se produce a través del sensor VirA-VirG que da como resultado la generación de una copia móvil de T-ADN monocatenario (cadena T). Una forma procesada de VirB2 es el componente principal del complejo T que se requiere para la transformación. VirD2 es la proteína que recubre el extremo 5′ de la cadena T transferida mediante unión covalente y se transporta al citoplasma de la célula hospedante. [8] [9] VirE2 es la proteína de unión al ADN monocatenario que presumiblemente recubre la cadena T en el citoplasma del hospedante mediante unión cooperativa . Luego se dirige al núcleo a través de interacciones con las proteínas de la célula huésped, como la importina a, la VirE3 bacteriana y proteínas similares a la dineína. Varios otros efectores de virulencia bacteriana como VirB5, VirB7 (los componentes menores del complejo T), VirD5, VirE2, VirE3 y VirF también pueden interactuar con las proteínas de las células vegetales huésped. [10]
La transferencia de T-ADN mediada por Agrobacterium se utiliza ampliamente como herramienta en biotecnología . Durante más de dos décadas, Agrobacterium tumefaciens se ha explotado para introducir genes en plantas para investigación básica, así como para la producción comercial de cultivos transgénicos . [11] En ingeniería genética , los genes promotores de tumores y de síntesis de opinas se eliminan del T-ADN y se reemplazan con un gen de interés y/o un marcador de selección, que se requiere para establecer qué plantas se han transformado con éxito. Los ejemplos de marcadores de selección incluyen neomicina fosfotransferasa, higromicina B fosfotransferasa (que fosforilan antibióticos) y fosfinotricina acetiltransferasa (que acetila y desactiva la fosfinotricina , un potente inhibidor de la glutamina sintetasa ) o formulaciones herbicidas como Basta o Bialophos. [12] Otro sistema de selección que se puede emplear es el uso de marcadores metabólicos como la fosfo-manosa isomerasa. [13] Agrobacterium se utiliza luego como vector para transferir el ADN-T diseñado a las células vegetales donde se integra en el genoma de la planta. Este método se puede utilizar para generar plantas transgénicas que porten un gen extraño. Agrobacterium tumefaciens es capaz de transferir ADN extraño tanto a plantas monocotiledóneas como dicotiledóneas de manera eficiente mientras se ocupa de factores de importancia crítica como el genotipo de las plantas, los tipos y edades de los tejidos inoculados, el tipo de vectores, las cepas de Agrobacterium , los genes marcadores de selección y los agentes selectivos, y varias condiciones de cultivo de tejidos. [4]
El mismo procedimiento de transferencia de T-ADN puede utilizarse para alterar genes mediante mutagénesis insercional . [6] La secuencia de T-ADN insertada no solo crea una mutación, sino que su inserción también "marca" [14] el gen afectado, lo que permite su aislamiento como secuencias flanqueantes de T-ADN. Un gen reportero puede vincularse al extremo derecho del T-ADN que se va a transformar junto con un replicón plasmídico y un gen de resistencia a antibióticos seleccionables (como la higromicina ) y puede explicitar aproximadamente el 30% de la eficiencia promedio de tener fusiones génicas inducidas por insertos de T-ADN exitosos en Arabidopsis thaliana . [15]
La genética inversa implica probar la función presunta de un gen que se conoce alterándolo y luego buscando el efecto de esa mutación inducida en el fenotipo del organismo. La mutagénesis por marcado de T-ADN implica la selección de poblaciones mediante mutaciones de inserción de T-ADN. Las colecciones de mutaciones de T-ADN conocidas proporcionan recursos para estudiar las funciones de genes individuales, como se desarrolló para la planta modelo Arabidopsis thaliana . [16] [17] Los ejemplos de mutaciones de inserción de T-ADN en Arabidopsis thaliana incluyen aquellos asociados con muchas clases de fenotipos, incluyendo plantas letales para plántulas, variantes de tamaño, variantes de pigmento, defectuosas para embriones, de fertilidad reducida y morfológica o fisiológicamente aberrantes. [18]