La reacción es análoga a la reacción catalizada por el citocromo bc 1 (complejo III) de la cadena de transporte de electrones mitocondrial . Durante la fotosíntesis , el complejo citocromo b 6 f es un paso a lo largo de la cadena que transfiere electrones del fotosistema II al fotosistema I y, al mismo tiempo, bombea protones al espacio tilacoide, lo que contribuye a la generación de un gradiente electroquímico (energético) [2] que luego se utiliza para sintetizar ATP a partir de ADP .
Estructura de la enzima
El complejo citocromo b 6 f es un dímero, y cada monómero está compuesto por ocho subunidades. [3] Estas consisten en cuatro subunidades grandes: un citocromo f de 32 kDa con un citocromo de tipo c, un citocromo b 6 de 25 kDa con un grupo hemo de potencial bajo y alto, una proteína de hierro-azufre de Rieske de 19 kDa que contiene un grupo [2Fe-2S] y una subunidad IV de 17 kDa; junto con cuatro subunidades pequeñas (3-4 kDa): PetG, PetL, PetM y PetN. [3] [4] El peso molecular total es de 217 kDa.
Se han determinado las estructuras cristalinas de los complejos de citocromo b 6 f de Chlamydomonas reinhardtii , Mastigocladus laminosus y Nostoc sp. PCC 7120. [2] [5] [6] [7] [8] [9]
El núcleo del complejo es estructuralmente similar al núcleo del citocromo bc 1. El citocromo b 6 y la subunidad IV son homólogos del citocromo b , [10] y las proteínas de hierro-azufre de Rieske de los dos complejos son homólogas. [11] Sin embargo, el citocromo f y el citocromo c 1 no son homólogos. [12]
El citocromo b 6 f contiene siete grupos prostéticos . [13] [14] Cuatro de ellos se encuentran tanto en el citocromo b 6 f como en el bc 1 : el hemo de tipo c del citocromo c 1 y f, los dos hemo de tipo b (b p y b n ) en bc 1 y b 6 f, y el grupo [2Fe-2S] de la proteína Rieske. Se encuentran tres grupos prostéticos únicos en el citocromo b 6 f: clorofila a , β-caroteno y hemo c n (también conocido como hemo x). [5]
El espacio intermonómero dentro del núcleo del dímero del complejo citocromo b6f está ocupado por lípidos, [9] lo que proporciona direccionalidad a la transferencia de electrones hemo-hemo a través de la modulación del entorno dieléctrico intraproteico. [15]
Función biológica
Mutante del citocromo b 6 f del tabaco ( Nicotiana tabacum ) (derecha) junto a una planta normal. Las plantas se utilizan en la investigación de la fotosíntesis para investigar la fotofosforilación cíclica.
En la fotosíntesis , el complejo citocromo b 6 f funciona para mediar la transferencia de electrones y de energía entre los dos complejos del centro de reacción fotosintético, el fotosistema II y el fotosistema I , mientras transfiere protones desde el estroma del cloroplasto a través de la membrana tilacoide hasta el lumen . [2] El transporte de electrones a través del citocromo b 6 f es responsable de crear el gradiente de protones que impulsa la síntesis de ATP en los cloroplastos. [4]
En una reacción separada, el complejo citocromo b 6 f juega un papel central en la fotofosforilación cíclica , cuando el NADP + no está disponible para aceptar electrones de la ferredoxina reducida . [16] Este ciclo, impulsado por la energía de P700 + , contribuye a la creación de un gradiente de protones que se puede utilizar para impulsar la síntesis de ATP. Se ha demostrado que este ciclo es esencial para la fotosíntesis, [17] ayudando a mantener la proporción adecuada de producción de ATP/NADPH para la fijación de carbono . [18] [19]
Las reacciones de desprotonación-oxidación de quinol del lado p dentro del complejo citocromo b6f se han implicado en la generación de especies reactivas de oxígeno. [20] Se ha sugerido que una molécula de clorofila integral ubicada dentro del sitio de oxidación de quinol realiza una función estructural, no fotoquímica, en la mejora de la tasa de formación de especies reactivas de oxígeno, posiblemente para proporcionar una vía redox para la comunicación intracelular. [21]
Mecanismo de reacción
El complejo citocromo b 6 f es responsable de la transferencia de electrones " no cíclica " (1) y " cíclica " (2) entre dos transportadores redox móviles, plastoquinol (QH 2 ) y plastocianina (Pc):
El citocromo b 6 f cataliza la transferencia de electrones del plastoquinol a la plastocianina, mientras bombea dos protones desde el estroma hacia el lumen del tilacoide:
Esta reacción ocurre a través del ciclo Q como en el Complejo III. [22] El plastoquinol actúa como transportador de electrones, transfiriendo sus dos electrones a cadenas de transporte de electrones (CTE) de alto y bajo potencial a través de un mecanismo llamado bifurcación de electrones . [23] El complejo contiene hasta tres moléculas de plastoquinona que forman una red de transferencia de electrones que son responsables del funcionamiento del ciclo Q y sus funciones de detección redox y catalíticas en la fotosíntesis. [24]
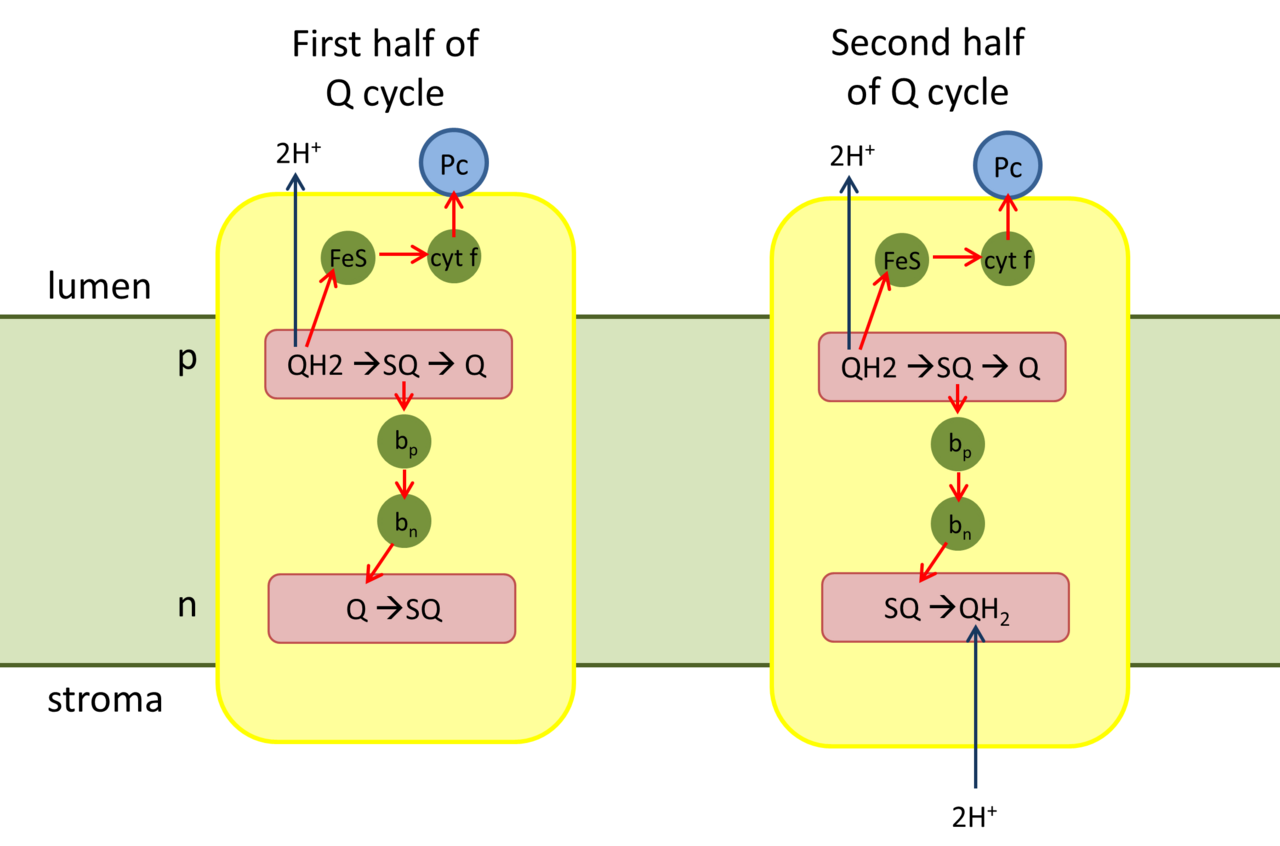
Ciclo Q
Ciclo Q del citocromo b 6 f
Primera mitad del ciclo Q
El QH 2 se une al lado "p" positivo (lado del lumen) del complejo. Se oxida a una semiquinona (SQ) por el centro de hierro-azufre (cadena de transporte de electrones de alto potencial) y libera dos protones al lumen del tilacoide [ cita requerida ] .
El centro de hierro-azufre reducido transfiere su electrón a través del citocromo f a Pc.
En la cadena de transporte de electrones de bajo potencial, SQ transfiere su electrón al hemo b p del citocromo b6.
Luego, el hemo b p transfiere el electrón al hemo b n .
El hemo b n reduce Q con un electrón para formar SQ.
Segunda mitad del ciclo Q
Un segundo QH 2 se une al complejo.
En la cadena de transporte de electrones de alto potencial, un electrón reduce otro Pc oxidado.
En la cadena de transporte de electrones de bajo potencial, el electrón del hemo b n se transfiere a SQ, y el Q 2− completamente reducido toma dos protones del estroma para formar QH 2 .
El Q oxidado y el QH 2 reducido que se ha regenerado se difunden en la membrana.
Transferencia cíclica de electrones
A diferencia del complejo III, el citocromo b 6 f cataliza otra reacción de transferencia de electrones que es fundamental para la fotofosforilación cíclica . El electrón de la ferredoxina (Fd) se transfiere a la plastoquinona y luego al complejo citocromo b 6 f para reducir la plastocianina, que es reoxidada por P700 en el fotosistema I. [25] El mecanismo exacto de la reducción de la plastoquinona por la ferredoxina aún está bajo investigación. Una propuesta es que existe una ferredoxina:plastoquinona-reductasa o una NADP deshidrogenasa. [25] Dado que el hemo x no parece ser necesario para el ciclo Q y no se encuentra en el complejo III, se ha propuesto que se utiliza para la fotofosforilación cíclica mediante el siguiente mecanismo: [23] [26]
Fd (rojo) + hemo x (buey) → Fd (buey) + hemo x (rojo)
^ abc Hasan SS, Yamashita E, Baniulis D, Cramer WA (marzo de 2013). "Vías de transferencia de protones dependientes de quinona en el complejo fotosintético del citocromo b6f". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 110 (11): 4297–302. doi : 10.1073/pnas.1222248110 . PMC 3600468 . PMID 23440205.
^ ab Whitelegge JP, Zhang H, Aguilera R, Taylor RM, Cramer WA (octubre de 2002). "Cromatografía líquida con espectrometría de masas de ionización por electrospray (LCMS+) de cobertura total de subunidades de un complejo de proteína de membrana oligomérica: citocromo b(6)f de espinaca y la cianobacteria Mastigocladus laminosus". Molecular & Cellular Proteomics . 1 (10): 816–27. doi : 10.1074/mcp.m200045-mcp200 . PMID 12438564.
^ ab Voet DJ, Voet JG (2011). Bioquímica . Nueva York, Nueva York: Wiley, J. ISBN978-0-470-57095-1.
^ ab Stroebel D, Choquet Y, Popot JL, Picot D (noviembre de 2003). "Un hemo atípico en el complejo citocromo b(6)f". Nature . 426 (6965): 413–8. doi :10.1038/nature02155. PMID 14647374. S2CID 130033.
^ Yamashita E, Zhang H, Cramer WA (junio de 2007). "Estructura del complejo citocromo b6f: inhibidores de análogos de quinona como ligandos del grupo hemo". Journal of Molecular Biology . 370 (1): 39–52. doi :10.1016/j.jmb.2007.04.011. PMC 1993820 . PMID 17498743.
^ Baniulis D, Yamashita E, Whitelegge JP, Zatsman AI, Hendrich MP, Hasan SS, Ryan CM, Cramer WA (abril de 2009). "Estructura-función, estabilidad y modificación química del complejo cianobacteriano citocromo b6f de Nostoc sp. PCC 7120". Revista de química biológica . 284 (15): 9861–9. doi : 10.1074/jbc.M809196200 . PMC 2665108 . PMID 19189962.
^ Hasan SS, Stofleth JT, Yamashita E, Cramer WA (abril de 2013). "Cambios conformacionales inducidos por lípidos dentro del complejo citocromo b6f de la fotosíntesis oxigénica". Bioquímica . 52 (15): 2649–54. doi :10.1021/bi301638h. PMC 4034689 . PMID 23514009.
^ ab Hasan SS, Cramer WA (julio de 2014). "Arquitectura lipídica interna del complejo hetero-oligomérico del citocromo b6f". Structure . 22 (7): 1008–15. doi :10.1016/j.str.2014.05.004. PMC 4105968 . PMID 24931468.
^ Widger WR, Cramer WA, Herrmann RG, Trebst A (febrero de 1984). "Homología de secuencia y similitud estructural entre el citocromo b del complejo mitocondrial III y el complejo b6-f del cloroplasto: posición de los hemo del citocromo b en la membrana". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 81 (3): 674–8. doi : 10.1073/pnas.81.3.674 . PMC 344897 . PMID 6322162.
^ Carrell CJ, Zhang H, Cramer WA, Smith JL (diciembre de 1997). "Identidad biológica y diversidad en la fotosíntesis y la respiración: estructura del dominio del lado del lumen de la proteína Rieske del cloroplasto". Estructura . 5 (12): 1613–25. doi : 10.1016/s0969-2126(97)00309-2 . PMID 9438861.
^ Martinez SE, Huang D, Szczepaniak A, Cramer WA, Smith JL (febrero de 1994). "La estructura cristalina del citocromo f del cloroplasto revela un nuevo pliegue del citocromo y una ligadura del hemo inesperada". Structure . 2 (2): 95–105. doi : 10.1016/s0969-2126(00)00012-5 . PMID 8081747.
^ Baniulis D, Yamashita E, Zhang H, Hasan SS, Cramer WA (2008). "Estructura-función del complejo citocromo b6f". Fotoquímica y fotobiología . 84 (6): 1349–58. doi :10.1111/j.1751-1097.2008.00444.x. PMID 19067956. S2CID 44992397.
^ Cramer WA, Zhang H, Yan J, Kurisu G, Smith JL (mayo de 2004). "Evolución de la fotosíntesis: estructura independiente del tiempo del complejo citocromo b6f". Bioquímica . 43 (20): 5921–9. doi :10.1021/bi049444o. PMID 15147175.
^ Hasan SS, Zakharov SD, Chauvet A, Stadnytskyi V, Savikhin S, Cramer WA (junio de 2014). "Un mapa de heterogeneidad dieléctrica en una proteína de membrana: el complejo hetero-oligomérico del citocromo b6f". The Journal of Physical Chemistry B . 118 (24): 6614–25. doi :10.1021/jp501165k. PMC 4067154 . PMID 24867491.
^ ab Berg JM, Tymoczko JL, Stryer L, Stryer L (2007). Bioquímica . Nueva York: WH Freeman. ISBN978-0-7167-8724-2.
^ Munekage Y, Hashimoto M, Miyake C, Tomizawa K, Endo T, Tasaka M, Shikanai T (junio de 2004). "El flujo cíclico de electrones alrededor del fotosistema I es esencial para la fotosíntesis". Nature . 429 (6991): 579–82. Bibcode :2004Natur.429..579M. doi :10.1038/nature02598. PMID 15175756. S2CID 4421776.
^ Blankenship RE (2002). Mecanismos moleculares de la fotosíntesis . Oxford; Malden, MA: Blackwell Science. ISBN978-0-632-04321-7.
^ Bendall D (1995). "Fotofosforilación cíclica y transporte de electrones". Biochimica et Biophysica Acta (BBA) - Bioenergetics . 1229 : 23–38. doi : 10.1016/0005-2728(94)00195-B .
^ Baniulis D, Hasan SS, Stofleth JT, Cramer WA (diciembre de 2013). "Mecanismo de producción mejorada de superóxido en el complejo citocromo b(6)f de la fotosíntesis oxigénica". Bioquímica . 52 (50): 8975–83. doi :10.1021/bi4013534. PMC 4037229 . PMID 24298890.
^ Hasan SS, Proctor EA, Yamashita E, Dokholyan NV, Cramer WA (octubre de 2014). "Tráfico dentro del complejo de lipoproteínas del citocromo b6f: activación del portal de quinonas". Revista biofísica . 107 (7): 1620–8. Código Bibliográfico :2014BpJ...107.1620H. doi :10.1016/j.bpj.2014.08.003. PMC 4190601 . PMID 25296314.
^ Cramer WA, Soriano GM, Ponomarev M, Huang D, Zhang H, Martinez SE, Smith JL (junio de 1996). "Algunos nuevos aspectos estructurales y antiguas controversias concernientes al complejo citocromo b6f de la fotosíntesis oxigénica". Revisión anual de fisiología vegetal y biología molecular de plantas . 47 : 477–508. doi :10.1146/annurev.arplant.47.1.477. PMID 15012298.
^ ab Cramer WA, Zhang H, Yan J, Kurisu G, Smith JL (2006). "Tráfico transmembrana en el complejo citocromo b6f". Revisión anual de bioquímica . 75 : 769–90. doi :10.1146/annurev.biochem.75.103004.142756. PMID 16756511.
^ Malone LA, Qian P, Mayneord GE, Hitchcock A, Farmer DA, Thompson RF, et al. (noviembre de 2019). "Estructura crio-EM del complejo citocromo B 6 F de la espinaca a una resolución de 3,6 Å" (PDF) . Nature . 575 (7783): 535–539. doi :10.1038/s41586-019-1746-6. PMID 31723268. S2CID 207987984.
^ ab Joliot P, Joliot A (julio de 2002). "Transferencia cíclica de electrones en hojas de plantas". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 99 (15): 10209–14. Bibcode :2002PNAS...9910209J. doi : 10.1073/pnas.102306999 . PMC 126649 . PMID 12119384.
^ Cramer WA, Yan J, Zhang H, Kurisu G, Smith JL (2005). "Estructura del complejo citocromo b6f: nuevos grupos prostéticos, espacio Q y la 'hipótesis de los entremeses' para el ensamblaje del complejo". Photosynthesis Research . 85 (1): 133–43. Bibcode :2005PhoRe..85..133C. doi :10.1007/s11120-004-2149-5. PMID 15977064. S2CID 20731696.
Lectura adicional
Sarewicz, M; Pintscher, S; Pietras, R; Borek, A; Bujnowicz, Ł; Hanke, G; Cramer, WA; Finazzi, G; Osyczka, A (24 de febrero de 2021). "Reacciones catalíticas y conservación de energía en los complejos de citocromo bc(1) y b(6)f de membranas transductoras de energía". Chemical Reviews . 121 (4): 2020–2108. doi : 10.1021/acs.chemrev.0c00712 . PMC 7908018 . PMID 33464892.
Enlaces externos
Estudios de estructura y función del complejo citocromo b6f: investigación actual sobre el citocromo b 6 f en el laboratorio de William Cramer en la Universidad de Purdue, EE. UU.
_cyt6bf_mutant.jpg/1280px-Tobacco_(Nicotiana_tabacum)_cyt6bf_mutant.jpg)

{kind=link}