Los murciélagos son mamíferos voladores del orden Chiroptera ( / k aɪ ˈ r ɒ p t ər ə / ). [a] Con sus extremidades anteriores adaptadas como alas , son los únicos mamíferos capaces de un vuelo verdadero y sostenido . Los murciélagos son más ágiles en vuelo que la mayoría de las aves, volando con sus dedos muy largos y extendidos cubiertos con una membrana delgada o patagio . El murciélago más pequeño, y posiblemente el mamífero existente más pequeño , es el murciélago hocico de cerdo de Kitti , que mide entre 29 y 34 milímetros ( 1+1 ⁄ 8 – 1+Miden entre 3 y 8 pulgadas de largo, 150 mm de ancho entre las alas y pesan entre22,6 g ( entre 1 y 3 onzas ) . Los murciélagos más grandes son los zorros voladores , y el zorro volador gigante de corona dorada ( Acerodon jubatus ) alcanza un peso de1,6 kg ( 3,5 libras ).+1 ⁄ 2 lb) y una envergadura de 1,7 m (5 pies 7 pulgadas).
El segundo orden más grande de mamíferos después de los roedores , los murciélagos comprenden alrededor del 20% de todas las especies de mamíferos clasificadas en todo el mundo, con más de 1.400 especies. Estos tradicionalmente se dividían en dos subórdenes: los megamurciélagos, principalmente frugívoros , y los micromurciélagos ecolocalizadores . Pero evidencia más reciente ha apoyado la división del orden en Yinpterochiroptera y Yangochiroptera , con megamurciélagos como miembros del primero junto con varias especies de micromurciélagos. Muchos murciélagos son insectívoros , y la mayoría del resto son frugívoros (comen fruta) o nectarívoros (comen néctar). Unas pocas especies se alimentan de animales distintos de los insectos; por ejemplo, los murciélagos vampiros se alimentan de sangre . La mayoría de los murciélagos son nocturnos , y muchos se posan en cuevas u otros refugios; no se sabe con certeza si los murciélagos tienen estos comportamientos para escapar de los depredadores . Los murciélagos están presentes en todo el mundo, con la excepción de las regiones extremadamente frías. Son importantes en sus ecosistemas para polinizar flores y dispersar semillas; muchas plantas tropicales dependen completamente de los murciélagos para estos servicios.
Los murciélagos proporcionan a los humanos algunos beneficios directos, a costa de algunas desventajas. El estiércol de murciélago se ha extraído como guano de cuevas y se ha utilizado como fertilizante. Los murciélagos consumen plagas de insectos, lo que reduce la necesidad de pesticidas y otras medidas de control de insectos. A veces son lo suficientemente numerosos y están lo suficientemente cerca de los asentamientos humanos como para servir como atracciones turísticas, y se utilizan como alimento en Asia y la Cuenca del Pacífico . Sin embargo, los murciélagos frugívoros a menudo son considerados plagas por los fruticultores. Debido a su fisiología, los murciélagos son un tipo de animal que actúa como reservorio natural de muchos patógenos , como la rabia ; y dado que son muy móviles, sociales y longevos, pueden propagar fácilmente enfermedades entre ellos. Si los humanos interactúan con los murciélagos, estos rasgos se vuelven potencialmente peligrosos para los humanos. Algunos murciélagos también son depredadores de mosquitos , suprimiendo la transmisión de enfermedades transmitidas por mosquitos .
Dependiendo de la cultura, los murciélagos pueden estar asociados simbólicamente con rasgos positivos, como la protección contra ciertas enfermedades o riesgos, el renacimiento o la larga vida, pero en Occidente, los murciélagos se asocian popularmente con la oscuridad, la malevolencia, la brujería, los vampiros y la muerte.
Un nombre inglés más antiguo para los murciélagos es flittermouse, que coincide con su nombre en otras lenguas germánicas (por ejemplo, el alemán Fledermaus y el sueco fladdermus ), relacionado con el aleteo de las alas. El inglés medio tenía bakke , muy probablemente cognado con el sueco antiguo natbakka ( ' murciélago nocturno ' ), que puede haber sufrido un cambio de -k- a -t- (al inglés moderno bat ) influenciado por el latín blatta , ' polilla, insecto nocturno ' . La palabra bat probablemente se utilizó por primera vez a principios de la década de 1570. [2] [3] El nombre Chiroptera deriva del griego antiguo : χείρ – cheir , ' mano ' [4] y πτερόν – pteron , ' ala ' . [1] [5]

Los delicados esqueletos de los murciélagos no se fosilizan bien; se estima que solo el 12% de los géneros de murciélagos que vivieron han sido encontrados en el registro fósil. [6] La mayoría de los fósiles de murciélagos más antiguos conocidos ya eran muy similares a los microquirópteros modernos, como Archaeopteropus (hace 32 millones de años). Los fósiles de murciélagos más antiguos conocidos incluyen Archaeonycteris praecursor y Altaynycteris aurora (hace 55-56 millones de años), ambos conocidos solo por dientes aislados. [7] [8] El esqueleto de murciélago completo más antiguo es Icaronycteris gunnelli (hace 52 millones de años), conocido a partir de dos esqueletos descubiertos en Wyoming. [9] [10] Los murciélagos extintos Palaeochiropteryx tupaiodon y Hassianycteris kumari , ambos de hace 48 millones de años, son los primeros mamíferos fósiles cuya coloración se ha descubierto: ambos eran de color marrón rojizo. [11] [12]
Los murciélagos se agrupaban anteriormente en el superorden Archonta , junto con las musarañas arbóreas (Scandentia), los colugos (Dermoptera) y los primates . [13] La evidencia genética moderna ahora ubica a los murciélagos en el superorden Laurasiatheria , con su taxón hermano como Ferungulata , que incluye a los carnívoros , pangolines , ungulados de dedos impares , ungulados de dedos pares y cetáceos . [14] [15] [16] [17] [18] Un estudio ubica a Chiroptera como un taxón hermano de los ungulados de dedos impares (Perissodactyla). [19]
La hipótesis de los primates voladores propuso que cuando se eliminan las adaptaciones al vuelo, los megamurciélagos se aliaron con los primates por características anatómicas que no comparten con los micromurciélagos y, por lo tanto, el vuelo evolucionó dos veces en los mamíferos. [20] Los estudios genéticos han apoyado firmemente la monofilia de los murciélagos y el origen único del vuelo de los mamíferos. [9] [20]
Un análisis molecular independiente que intentaba establecer las fechas en las que evolucionaron los ectoparásitos de murciélagos ( chinches ) llegó a la conclusión de que las chinches similares a las que conocemos hoy (todos los linajes principales existentes, todos los cuales se alimentan principalmente de murciélagos) ya se habían diversificado y establecido hace más de 100 millones de años (es decir, mucho antes de los registros más antiguos de murciélagos, 52 millones de años), lo que sugiere que inicialmente todos evolucionaron en huéspedes que no eran murciélagos y "los murciélagos fueron colonizados varias veces de forma independiente, a menos que el origen evolutivo de los murciélagos haya sido subestimado groseramente". [21] Las pulgas , como grupo, son bastante antiguas (la mayoría de las familias de pulgas se formaron alrededor del final del Cretácico [22] ), pero ningún análisis ha proporcionado estimaciones de la edad de los linajes de pulgas asociados con los murciélagos. Sin embargo, los miembros más antiguos conocidos de un linaje diferente de ectoparásitos de murciélagos ( moscas murciélago ) son de aproximadamente 20 millones de años, mucho después del origen de los murciélagos. [23] La familia de tijeretas ectoparásitas de murciélagos Arixeniidae no tiene registro fósil, pero no se cree que se originó hace más de 23 millones de años. [24]
La evidencia genética indica que los megamurciélagos se originaron durante el Eoceno temprano y pertenecen a las cuatro líneas principales de micromurciélagos. [18] Se han propuesto dos nuevos subórdenes; Yinpterochiroptera incluye la familia Pteropodidae , o megamurciélagos, así como las familias Rhinolophidae , Hipposideridae , Craseonycteridae , Megadermatidae y Rhinopomatidae . [26] Yangochiroptera incluye las otras familias de murciélagos (todas las cuales utilizan la ecolocalización laríngea), una conclusión respaldada por un estudio de ADN de 2005. [26] Un estudio filogenómico de 2013 respaldó los dos nuevos subórdenes propuestos. [18]

El descubrimiento en 2003 de un fósil de murciélago temprano de la Formación Green River de 52 millones de años , Onychonycteris finneyi , indica que el vuelo evolucionó antes que las habilidades ecolocativas. [27] [28] Onychonycteris tenía garras en sus cinco dedos, mientras que los murciélagos modernos tienen como máximo dos garras en dos dígitos de cada mano. También tenía patas traseras más largas y antebrazos más cortos, similares a los mamíferos trepadores que se cuelgan de las ramas, como los perezosos y los gibones . Este murciélago del tamaño de la palma de la mano tenía alas cortas y anchas, lo que sugiere que no podía volar tan rápido ni tan lejos como las especies de murciélagos posteriores. En lugar de batir sus alas continuamente mientras volaba, Onychonycteris probablemente alternaba entre aleteos y planeos en el aire. [9] Esto sugiere que este murciélago no volaba tanto como los murciélagos modernos, sino que volaba de árbol en árbol y pasaba la mayor parte del tiempo trepando o colgando de las ramas. [29] Las características distintivas del fósil de Onychonycteris también respaldan la hipótesis de que el vuelo de los mamíferos probablemente evolucionó en locomotoras arbóreas, en lugar de corredores terrestres. Este modelo de desarrollo del vuelo, conocido comúnmente como la teoría de los "árboles caídos", sostiene que los murciélagos volaron primero aprovechando la altura y la gravedad para caer sobre sus presas, en lugar de correr lo suficientemente rápido como para despegar a nivel del suelo. [30] [31]
La filogenia molecular fue controvertida, ya que apuntaba a que los micro murciélagos no tenían un ancestro común único , lo que implicaba que ocurrieron algunas transformaciones aparentemente improbables. La primera es que la ecolocalización laríngea evolucionó dos veces en los murciélagos, una en Yangochiroptera y otra en los rinolofoides. [32] La segunda es que la ecolocalización laríngea tuvo un único origen en Chiroptera, se perdió posteriormente en la familia Pteropodidae (todos los mega murciélagos) y luego evolucionó como un sistema de chasquido de lengua en el género Rousettus . [33] Los análisis de la secuencia del gen de vocalización FoxP2 no fueron concluyentes sobre si la ecolocalización laríngea se perdió en los pteropódidos o se ganó en los linajes ecolocalizadores. [34] La ecolocalización probablemente se derivó por primera vez en los murciélagos de llamadas comunicativas. Los murciélagos del Eoceno Icaronycteris (hace 52 millones de años) y Palaeochiropteryx tenían adaptaciones craneales que sugieren una capacidad para detectar ultrasonidos . Es posible que al principio se utilizara principalmente para buscar insectos en el suelo y mapear sus alrededores en su fase de planeo, o con fines comunicativos. Después de que se estableció la adaptación del vuelo, es posible que se haya perfeccionado para localizar presas voladoras mediante ecolocalización. [29] Los análisis del gen de la audición Prestin parecen favorecer la idea de que la ecolocalización se desarrolló de forma independiente al menos dos veces, en lugar de perderse de forma secundaria en los pteropódidos, [35] pero el análisis ontogénico de la cóclea respalda que la ecolocalización laríngea evolucionó solo una vez. [36]
Los murciélagos son mamíferos placentarios . Después de los roedores , son el orden más grande , representando aproximadamente el 20% de las especies de mamíferos. [37] En 1758, Carl Linnaeus clasificó las siete especies de murciélagos que conocía en el género Vespertilio en el orden Primates . Unos veinte años después, el naturalista alemán Johann Friedrich Blumenbach les dio su propio orden, Chiroptera. [38] Desde entonces, el número de especies descritas ha aumentado a más de 1.400, [39] tradicionalmente clasificadas en dos subórdenes: Megachiroptera (megamurciélagos) y Microchiroptera (micromurciélagos/murciélagos ecolocalizadores). [40] No todos los megamurciélagos son más grandes que los micromurciélagos. [41] Varias características distinguen a los dos grupos. Los micromurciélagos usan la ecolocalización para navegar y encontrar presas, pero los megamurciélagos aparte de los del género Rousettus no lo hacen. [42] En consecuencia, los megamurciélagos tienen una vista bien desarrollada. [40] Los megamurciélagos tienen una garra en el segundo dedo de la extremidad anterior. [43] [44] Las orejas externas de los micromurciélagos no se cierran para formar un anillo; los bordes están separados entre sí en la base de la oreja. [44] Los megamurciélagos comen fruta , néctar o polen, mientras que la mayoría de los micromurciélagos comen insectos ; otros se alimentan de fruta, néctar, polen, peces , ranas, pequeños mamíferos o sangre . [40]

A continuación se muestra un cuadro de tabla que sigue la clasificación de las familias de murciélagos reconocidas por varios autores del noveno volumen del Manual de los Mamíferos del Mundo publicado en 2019: [45]

La forma de la cabeza y los dientes de los murciélagos puede variar según la especie. En general, los megamurciélagos tienen hocicos más largos, cuencas oculares más grandes y orejas más pequeñas, lo que les da una apariencia más parecida a la de un perro, de ahí su apodo de "zorros voladores". [46] Entre los micromurciélagos, los hocicos más largos se asocian con la alimentación de néctar. [47] mientras que los murciélagos vampiros tienen hocicos reducidos para acomodar incisivos y caninos grandes. [48]
Los murciélagos insectívoros pequeños pueden tener hasta 38 dientes, mientras que los murciélagos vampiros tienen solo 20. Los murciélagos que se alimentan de insectos de caparazón duro tienen menos dientes pero más grandes con caninos más largos y mandíbulas inferiores más robustas que las especies que se alimentan de insectos de cuerpo más blando. En los murciélagos que se alimentan de néctar, los caninos son largos mientras que los molares son reducidos. En los murciélagos frugívoros, las cúspides de los molares están adaptadas para triturar. [47] Los incisivos superiores de los murciélagos vampiros carecen de esmalte , lo que los mantiene afilados como navajas. [48] La fuerza de mordida de los murciélagos pequeños se genera a través de la ventaja mecánica , lo que les permite morder a través de la armadura endurecida de los insectos o la piel de la fruta. [49]
Los murciélagos son los únicos mamíferos capaces de realizar un vuelo sostenido, a diferencia del planeo , como en el caso de la ardilla voladora . [50] El murciélago más rápido, el murciélago de cola libre mexicano ( Tadarida brasiliensis ), puede alcanzar una velocidad terrestre de 160 km/h (100 mph). [51]
Los huesos de los dedos de los murciélagos son mucho más flexibles que los de otros mamíferos, debido a su sección transversal aplanada y a los bajos niveles de calcio cerca de sus puntas. [52] [53] El alargamiento de los dígitos de los murciélagos, una característica clave necesaria para el desarrollo de las alas, se debe a la regulación positiva de las proteínas morfogenéticas óseas (Bmps). Durante el desarrollo embrionario , el gen que controla la señalización de Bmp, Bmp2 , está sujeto a una mayor expresión en las extremidades anteriores de los murciélagos, lo que resulta en la extensión de los dígitos manuales. Esta alteración genética crucial ayuda a crear las extremidades especializadas necesarias para el vuelo propulsado. La proporción relativa de dígitos de las extremidades anteriores de los murciélagos existentes en comparación con los de los murciélagos fósiles del Eoceno no tiene diferencias significativas, lo que sugiere que la morfología de las alas de los murciélagos se ha conservado durante más de cincuenta millones de años. [54] Durante el vuelo, los huesos sufren tensiones de flexión y cizallamiento ; las tensiones de flexión que se sienten son menores que en los mamíferos terrestres, pero la tensión de cizallamiento es mayor. Los huesos de las alas de los murciélagos tienen un punto de tensión de rotura ligeramente menor que los de las aves. [55]
Al igual que en otros mamíferos, y a diferencia de las aves, el radio es el componente principal del antebrazo. Los murciélagos tienen cinco dedos alargados, que irradian alrededor de la muñeca. El pulgar apunta hacia adelante y sostiene el borde delantero del ala, y los otros dedos sostienen la tensión que se mantiene en la membrana del ala. El segundo y tercer dedo van a lo largo de la punta del ala, lo que permite que el ala se tire hacia adelante contra la resistencia aerodinámica , sin tener que ser gruesa como en las alas de los pterosaurios . El cuarto y quinto dedo van desde la muñeca hasta el borde posterior , y repelen la fuerza de flexión causada por el aire que empuja contra la membrana rígida. [56] Debido a sus articulaciones flexibles, los murciélagos son más maniobrables y más diestros que los mamíferos planeadores. [57]

Las alas de los murciélagos son mucho más delgadas y constan de más huesos que las alas de las aves, lo que les permite maniobrar con mayor precisión que estos últimos y volar con más sustentación y menos resistencia. [58] Al plegar las alas hacia sus cuerpos en la carrera ascendente, ahorran un 35 por ciento de energía durante el vuelo. [59] Las membranas son delicadas y se desgarran fácilmente, [60] pero pueden volver a crecer y los pequeños desgarros se curan rápidamente. [60] [61] La superficie de las alas está equipada con receptores sensibles al tacto en pequeñas protuberancias llamadas células de Merkel , que también se encuentran en las yemas de los dedos humanos. Estas áreas sensibles son diferentes en los murciélagos, ya que cada protuberancia tiene un pequeño pelo en el centro, lo que la hace aún más sensible y permite al murciélago detectar y adaptarse a los cambios en el flujo de aire; el uso principal es juzgar la velocidad más eficiente a la que volar, y posiblemente también para evitar paradas . [62] Los murciélagos insectívoros también pueden usar pelos táctiles para ayudar a realizar maniobras complejas para capturar presas en vuelo. [57]
El patagio es la membrana del ala; se extiende entre los huesos del brazo y los dedos, y baja por el costado del cuerpo hasta las extremidades traseras y la cola. Esta membrana de la piel consta de tejido conectivo , fibras elásticas , nervios , músculos y vasos sanguíneos . Los músculos mantienen la membrana tensa durante el vuelo. [63] El grado en que la cola de un murciélago está unida a un patagio puede variar según la especie, y algunos tienen colas completamente libres o incluso ninguna cola. [47] La piel del cuerpo del murciélago, que tiene una capa de epidermis y dermis , así como folículos pilosos , glándulas sudoríparas y una capa subcutánea grasa, es muy diferente de la piel de la membrana del ala. Dependiendo de la especie de murciélago, la presencia de folículos pilosos y glándulas sudoríparas variará en el patagio. [64] Este patagio es una doble capa extremadamente delgada de epidermis; estas capas están separadas por un centro de tejido conectivo , rico en colágeno y fibras elásticas . En algunas especies de murciélagos, las glándulas sudoríparas estarán presentes entre este tejido conectivo. [65] Además, si hay folículos pilosos, esto ayuda al murciélago a ajustar las maniobras de vuelo repentinas. [66] [67] Para los embriones de murciélago, la apoptosis (muerte celular programada) afecta solo a las extremidades traseras, mientras que las extremidades delanteras retienen membranas entre los dedos que forman las membranas de las alas. [68] A diferencia de las aves, cuyas alas rígidas producen tensión de flexión y torsión en los hombros, los murciélagos tienen una membrana alar flexible que puede resistir solo la tensión. Para lograr volar, un murciélago ejerce fuerza hacia adentro en los puntos donde la membrana se encuentra con el esqueleto, de modo que una fuerza opuesta la equilibra en los bordes del ala perpendiculares a la superficie del ala. Esta adaptación no permite a los murciélagos reducir su envergadura, a diferencia de las aves, que pueden plegar parcialmente sus alas en vuelo, reduciendo radicalmente la envergadura y el área del ala para el movimiento ascendente y para el planeo. Por lo tanto, los murciélagos no pueden viajar largas distancias como lo hacen las aves. [56]
Los murciélagos que se alimentan de néctar y polen pueden flotar, de manera similar a los colibríes . Los bordes afilados de sus alas pueden crear vórtices que les proporcionan sustentación . El vórtice puede estabilizarse si el animal modifica la curvatura de sus alas. [69]

Cuando no vuelan, los murciélagos cuelgan boca abajo de sus patas, una postura conocida como percha. [70] Los fémures están unidos a las caderas de una manera que les permite doblarse hacia afuera y hacia arriba en el vuelo. La articulación del tobillo puede flexionarse para permitir que el borde posterior de las alas se doble hacia abajo. Esto no les permite muchos movimientos aparte de colgarse o trepar a los árboles. [56] La mayoría de los megamurciélagos se posan con la cabeza metida hacia el vientre, mientras que la mayoría de los micromurciélagos se posan con el cuello enroscado hacia la espalda. Esta diferencia se refleja en la estructura de las vértebras cervicales o del cuello en los dos grupos, que son claramente distintas. [70] Los tendones permiten a los murciélagos bloquear sus patas cerradas cuando cuelgan de un percha. Se necesita fuerza muscular para soltarse, pero no para agarrar una percha o para sujetarse. [71]
Cuando están en el suelo, la mayoría de los murciélagos sólo pueden arrastrarse torpemente. Unas pocas especies, como el murciélago de cola corta menor de Nueva Zelanda y el murciélago vampiro común, son ágiles en el suelo. Ambas especies realizan marchas laterales (las extremidades se mueven una tras otra) cuando se desplazan lentamente, pero los murciélagos vampiro se mueven con una marcha a saltos (todas las extremidades se mueven al unísono) a mayor velocidad, utilizando las alas plegadas para impulsarse hacia delante. Los murciélagos vampiros probablemente desarrollaron estas marchas para seguir a sus anfitriones, mientras que los murciélagos de cola corta se desarrollaron en ausencia de competidores mamíferos terrestres. La locomoción terrestre mejorada no parece haber reducido su capacidad de volar. [72]
Los murciélagos tienen un sistema circulatorio eficiente . Parecen hacer uso de una venomoción particularmente fuerte, una contracción rítmica de los músculos de la pared venosa . En la mayoría de los mamíferos, las paredes de las venas proporcionan principalmente resistencia pasiva, manteniendo su forma a medida que fluye sangre desoxigenada a través de ellas, pero en los murciélagos parecen apoyar activamente el flujo sanguíneo de regreso al corazón con esta acción de bombeo. [73] [74] Dado que sus cuerpos son relativamente pequeños y livianos, los murciélagos no corren el riesgo de que el flujo sanguíneo se les suba a la cabeza cuando están descansando. [75]
Los murciélagos poseen un sistema respiratorio altamente adaptado para hacer frente a las exigencias del vuelo propulsado, una actividad que exige mucho esfuerzo energético y un gran flujo continuo de oxígeno. En los murciélagos, la superficie alveolar relativa y el volumen sanguíneo capilar pulmonar son mayores que en la mayoría de los demás mamíferos cuadrúpedos pequeños. [76] Durante el vuelo, el ciclo respiratorio tiene una relación de uno a uno con el ciclo de aleteo. [77] Debido a las limitaciones de los pulmones de los mamíferos, los murciélagos no pueden mantener un vuelo a gran altitud. [56]
.jpg/1280px-Flying_fox_at_botanical_gardens_in_Sydney_(cropped).jpg)
Para ejercitar los músculos de vuelo de los murciélagos se necesita mucha energía y un sistema circulatorio eficiente. El suministro de energía a los músculos que participan en el vuelo requiere aproximadamente el doble de cantidad en comparación con los músculos que no utilizan el vuelo como medio de locomoción de los mamíferos. Paralelamente al consumo de energía, los niveles de oxígeno en sangre de los animales voladores son el doble que los de los mamíferos que se desplazan terrestres. Como el suministro de sangre controla la cantidad de oxígeno suministrado a todo el cuerpo, el sistema circulatorio debe responder en consecuencia. Por lo tanto, en comparación con un mamífero terrestre del mismo tamaño relativo, el corazón del murciélago puede ser hasta tres veces más grande y bombear más sangre. [79] El gasto cardíaco se deriva directamente de la frecuencia cardíaca y el volumen sistólico de la sangre; [80] un micromurciélago activo puede alcanzar una frecuencia cardíaca de 1000 latidos por minuto . [81]
Gracias a su tejido membranoso extremadamente fino, las alas de un murciélago pueden contribuir significativamente a la eficiencia total del intercambio de gases del organismo. [82] Debido a la alta demanda energética del vuelo, el cuerpo del murciélago satisface esas demandas intercambiando gases a través del patagio del ala. Cuando el murciélago tiene las alas extendidas, permite un aumento en la relación entre el área de superficie y el volumen. El área de superficie de las alas es aproximadamente el 85% de la superficie corporal total, lo que sugiere la posibilidad de un grado útil de intercambio de gases. [82] Los vasos subcutáneos en la membrana se encuentran muy cerca de la superficie y permiten la difusión de oxígeno y dióxido de carbono. [83]
El sistema digestivo de los murciélagos tiene distintas adaptaciones dependiendo de la especie de murciélago y su dieta. Como en otros animales voladores, la comida se procesa de forma rápida y eficaz para satisfacer la demanda energética. Los murciélagos insectívoros pueden tener ciertas enzimas digestivas para procesar mejor a los insectos, como la quitinasa para descomponer la quitina , que es un gran componente de los insectos. [84] Los murciélagos vampiros, probablemente debido a su dieta de sangre, son los únicos vertebrados que no tienen la enzima maltasa , que descompone el azúcar de malta , en su tracto intestinal. Los murciélagos néctivoros y frugívoros tienen más enzimas maltasa y sacarasa que los insectívoros, para hacer frente a los mayores contenidos de azúcar de su dieta. [85]
Las adaptaciones de los riñones de los murciélagos varían con sus dietas. Los murciélagos carnívoros y vampiros consumen grandes cantidades de proteínas y pueden producir orina concentrada ; sus riñones tienen una corteza delgada y papilas renales largas . Los murciélagos frugívoros carecen de esa capacidad y tienen riñones adaptados para la retención de electrolitos debido a su dieta baja en electrolitos; en consecuencia, sus riñones tienen una corteza gruesa y papilas cónicas muy cortas. [85] Los murciélagos tienen tasas metabólicas más altas asociadas con el vuelo, lo que conduce a una mayor pérdida de agua respiratoria. Sus grandes alas están compuestas de membranas altamente vascularizadas, lo que aumenta el área de superficie y conduce a la pérdida de agua por evaporación cutánea . [78] El agua ayuda a mantener su equilibrio iónico en la sangre, el sistema de termorregulación y la eliminación de desechos y toxinas del cuerpo a través de la orina. También son susceptibles a la intoxicación por urea en la sangre si no reciben suficiente líquido. [86]
La estructura del sistema uterino en los murciélagos hembras puede variar según la especie: algunas tienen dos cuernos uterinos mientras que otras tienen una sola cámara principal. [87]
Los micro murciélagos y algunos mega murciélagos emiten sonidos ultrasónicos para producir ecos. La intensidad del sonido de estos ecos depende de la presión subglótica. El músculo cricotiroideo de los murciélagos controla la frecuencia del pulso de orientación, que es una función importante. Este músculo está ubicado dentro de la laringe y es el único músculo tensor capaz de ayudar a la fonación. [88] Al comparar el pulso saliente con los ecos de retorno, los murciélagos pueden recopilar información sobre su entorno. Esto les permite detectar presas en la oscuridad. [89] Algunos llamados de murciélagos pueden alcanzar los 140 decibeles . [90] Los micro murciélagos usan su laringe para emitir señales de ecolocalización a través de la boca o la nariz. [91] Los llamados de micro murciélagos varían en frecuencia de 14.000 a más de 100.000 Hz, extendiéndose mucho más allá del rango de audición humana (entre 20 y 20.000 Hz). [92] Varios grupos de murciélagos han desarrollado extensiones carnosas alrededor y por encima de las fosas nasales, conocidas como hojas nasales , que desempeñan un papel en la transmisión del sonido. [93]
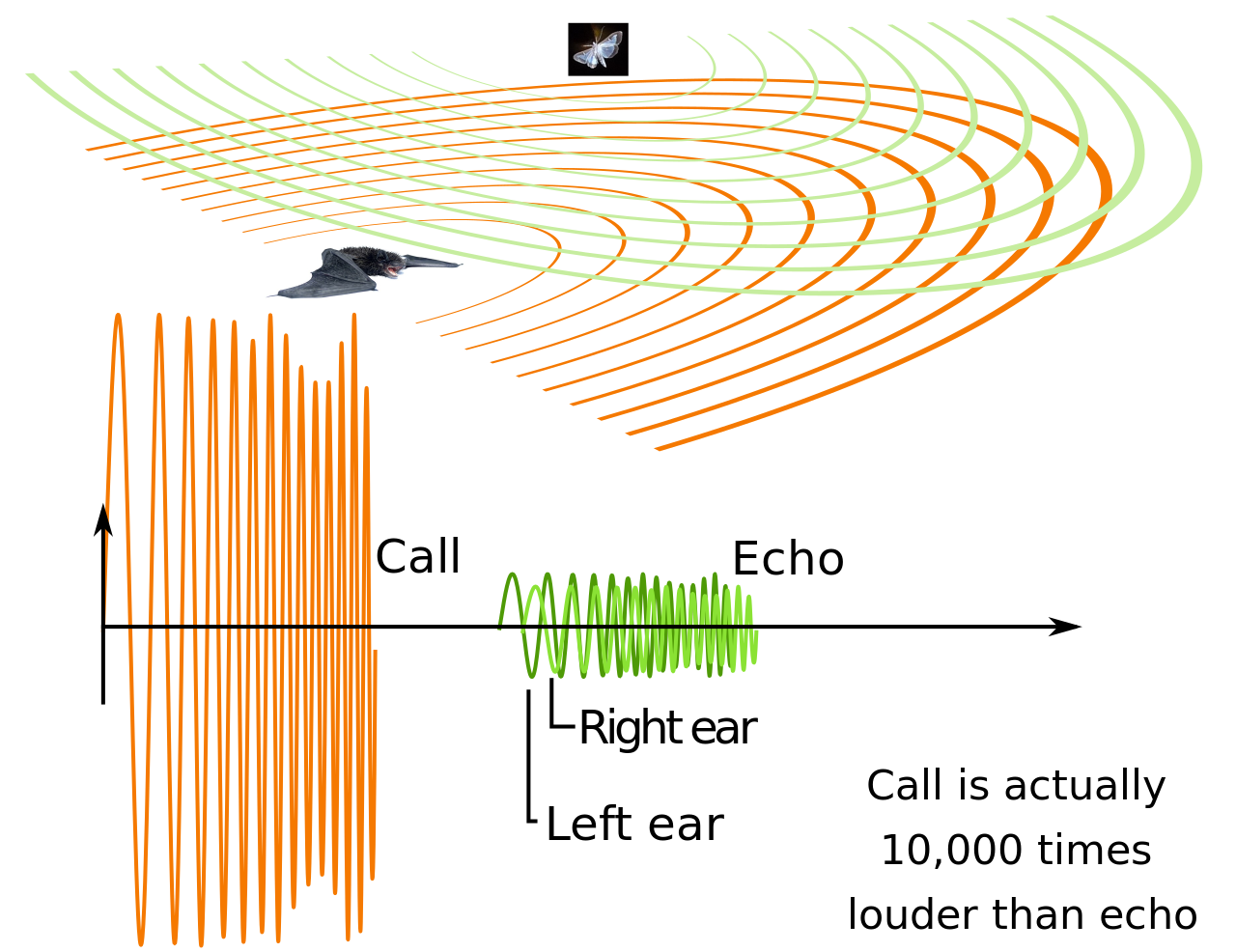
En la ecolocalización de ciclo de trabajo bajo, los murciélagos pueden separar sus llamadas y los ecos que regresan por tiempo. Tienen que cronometrar sus llamadas cortas para que terminen antes de que regresen los ecos. [94] El retraso de los ecos de retorno permite al murciélago estimar la distancia a su presa. [92] En la ecolocalización de ciclo de trabajo alto, los murciélagos emiten una llamada continua y separan el pulso y el eco en frecuencia utilizando el efecto Doppler de su movimiento en vuelo. El cambio de los ecos de retorno proporciona información relacionada con el movimiento y la ubicación de la presa del murciélago. Estos murciélagos deben lidiar con los cambios en el efecto Doppler debido a los cambios en su velocidad de vuelo. Se han adaptado a cambiar su frecuencia de emisión de pulsos en relación con su velocidad de vuelo para que los ecos aún regresen en el rango de audición óptimo. [94] [95]
Además de ecolocalizar a sus presas, las orejas de los murciélagos son sensibles a los sonidos que emiten sus presas, como el aleteo de las alas de las polillas. La compleja geometría de las crestas en la superficie interna de las orejas de los murciélagos ayuda a enfocar nítidamente las señales de ecolocalización y a escuchar pasivamente cualquier otro sonido producido por la presa. Estas crestas pueden considerarse el equivalente acústico de una lente de Fresnel y existen en una gran variedad de animales no relacionados, como el aye-aye , el gálago menor , el zorro orejudo , el lémur ratón y otros. [96] [97] [98] Los murciélagos pueden estimar la elevación de su objetivo utilizando los patrones de interferencia de los ecos que se reflejan en el trago , un colgajo de piel en el oído externo. [92]

Mediante el escaneo repetido, los murciélagos pueden construir mentalmente una imagen precisa del entorno en el que se mueven y de sus presas. [101] Algunas especies de polillas han explotado esto, como las polillas tigre , que producen señales de ultrasonido aposemáticas para advertir a los murciélagos que están químicamente protegidas y, por lo tanto, son desagradables. [99] [100] Las especies de polillas, incluida la polilla tigre, pueden producir señales para bloquear la ecolocalización de los murciélagos . Muchas especies de polillas tienen un órgano auditivo llamado tímpano , que responde a una señal entrante de murciélago haciendo que los músculos de vuelo de la polilla se contraigan erráticamente, enviando a la polilla a maniobras evasivas aleatorias. [102] [103] [104]
Los ojos de la mayoría de las especies de microquirópteros son pequeños y poco desarrollados, lo que da lugar a una agudeza visual deficiente , pero ninguna especie es ciega. [105] La mayoría de los microquirópteros tienen visión mesópica , lo que significa que pueden detectar luz solo en niveles bajos, mientras que otros mamíferos tienen visión fotópica , que les permite la visión en color. Los microquirópteros pueden utilizar su visión para orientarse y mientras viajan entre sus zonas de descanso y alimentación, ya que la ecolocalización es efectiva solo en distancias cortas. Algunas especies pueden detectar la luz ultravioleta (UV). Como los cuerpos de algunos microquirópteros tienen una coloración distinta, pueden ser capaces de discriminar colores. [50] [106] [107] [108]
Las especies de murciélagos gigantes suelen tener una vista tan buena como la humana, si no mejor. Su vista está adaptada tanto a la visión nocturna como a la diurna, incluida cierta visión en color. [108]
Los microquirópteros hacen uso de la magnetorrecepción , ya que tienen una alta sensibilidad al campo magnético de la Tierra , como lo hacen las aves. Los microquirópteros utilizan una brújula basada en la polaridad, lo que significa que diferencian el norte del sur, a diferencia de las aves, que utilizan la fuerza del campo magnético para diferenciar las latitudes , que pueden usarse en viajes de larga distancia. El mecanismo es desconocido, pero puede involucrar partículas de magnetita . [109] [110]

La mayoría de los murciélagos son homeotermicos (tienen una temperatura corporal estable), con la excepción de los murciélagos vespertinos (Vespertilionidae), los murciélagos de herradura (Rhinolophidae), los murciélagos de cola libre (Molossidae) y los murciélagos de alas dobladas (Miniopteridae), que utilizan ampliamente la heterotermia (donde la temperatura corporal puede variar). [111] [112] En comparación con otros mamíferos, los murciélagos tienen una alta conductividad térmica . Las alas están llenas de vasos sanguíneos y pierden calor corporal cuando se extienden. En reposo, pueden envolver sus alas alrededor de sí mismos para atrapar una capa de aire cálido. Los murciélagos más pequeños generalmente tienen una tasa metabólica más alta que los murciélagos más grandes y, por lo tanto, necesitan consumir más alimentos para mantener la homeotermia. [113]
Los murciélagos pueden evitar volar durante el día para prevenir el sobrecalentamiento por el sol, ya que sus membranas oscuras en las alas absorben la radiación solar. Los murciélagos pueden no ser capaces de disipar el calor si la temperatura ambiente es demasiado alta; [114] utilizan la saliva para refrescarse en condiciones extremas. [56] Entre los megamurciélagos, el zorro volador Pteropus hypomelanus utiliza la saliva y el abanicado de sus alas para refrescarse mientras se posa durante la parte más calurosa del día. [115] Entre los micromurciélagos, el murciélago de Yuma ( Myotis yumanensis ), el murciélago de cola libre mexicano y el murciélago pálido ( Antrozous pallidus ) hacen frente a temperaturas de hasta 45 °C (113 °F) jadeando, salivando y lamiendo su pelaje para promover el enfriamiento por evaporación; esto es suficiente para disipar el doble de su producción de calor metabólico. [116]
Los murciélagos también poseen un sistema de válvulas de esfínter en el lado arterial de la red vascular que corre a lo largo del borde de sus alas. Cuando están completamente abiertas, permiten que la sangre oxigenada fluya a través de la red capilar a través de la membrana del ala; cuando están contraídas, desvían el flujo directamente a las venas, evitando los capilares del ala. Esto permite a los murciélagos controlar la cantidad de calor que se intercambia a través de la membrana de vuelo, lo que les permite liberar calor durante el vuelo. Muchos otros mamíferos utilizan la red capilar en las orejas de gran tamaño con el mismo propósito. [117]

El letargo , un estado de actividad reducida en el que la temperatura corporal y el metabolismo disminuyen, es especialmente útil para los murciélagos, ya que utilizan una gran cantidad de energía mientras están activos, dependen de una fuente de alimento poco fiable y tienen una capacidad limitada para almacenar grasa. Por lo general, bajan su temperatura corporal en este estado a 6-30 °C (43-86 °F) y pueden reducir su gasto de energía entre un 50 y un 99 %. [118] Los murciélagos tropicales pueden usarlo para evitar la depredación, al reducir la cantidad de tiempo que pasan buscando alimento y, por lo tanto, reduciendo la posibilidad de ser atrapados por un depredador. [119] En general, se creía que los megamurciélagos eran homeotérmicos, pero se han identificado tres especies de megamurciélagos pequeños, con una masa de unos 50 gramos ( 1+Se sabe que los murciélagos de las flores (Syconycteris australis), el murciélago nectarívoro de lengua larga (Macroglossus minimus) y el murciélago de nariz tubular oriental (Nyctimene robinsoni) utilizan el letargo . Los estados de letargo duran más en verano para los megamurciélagos que en invierno. [120]
Durante la hibernación , los murciélagos entran en un estado tórpido y disminuyen su temperatura corporal durante el 99,6% de su período de hibernación; incluso durante los períodos de excitación, cuando su temperatura corporal vuelve a la normalidad, a veces entran en un estado tórpido superficial, conocido como "excitación heterotérmica". [121] Algunos murciélagos se vuelven inactivos durante las temperaturas más altas para mantenerse frescos en los meses de verano. [122]
Los murciélagos heterotérmicos durante largas migraciones pueden volar de noche y entrar en un estado de letargo, donde se posan durante el día. A diferencia de las aves migratorias, que vuelan durante el día y se alimentan durante la noche, los murciélagos nocturnos tienen un conflicto entre viajar y comer. La energía ahorrada reduce su necesidad de alimentarse y también disminuye la duración de la migración, lo que puede evitar que pasen demasiado tiempo en lugares desconocidos y disminuir la depredación. En algunas especies, las hembras preñadas pueden no entrar en estado de letargo. [123] [124]
El murciélago más pequeño es el murciélago nariz de cerdo de Kitti ( Craseonycteris thonglongyai ), que mide entre 29 y 34 mm ( 1+1 ⁄ 8 – 1+Mide aproximadamente 3 ⁄ 8 pulgadas de largo con una envergadura de 150 milímetros (6 pulgadas) y pesa entre2 y 2,6 g ( 1 ⁄ 16 y 3 ⁄ 32 oz). [125] También podría decirse que es la especie de mamífero existente más pequeña , junto con la musaraña etrusca . [126] Los murciélagos más grandes son algunas especies demegabats Pteropus y el zorro volador gigante de corona dorada ( Acerodon jubatus ), que puede pesar1,6 kg ( 3+1 ⁄ 2 lb) con una envergadura de 1,7 m (5 pies 7 pulgadas). [127] Los murciélagos más grandes tienden a utilizar frecuencias más bajas y los murciélagos más pequeños, frecuencias más altas para la ecolocalización; la ecolocalización de alta frecuencia es mejor para detectar presas más pequeñas. Las presas pequeñas pueden estar ausentes en las dietas de los murciélagos grandes, ya que no pueden detectarlas. [128] Las adaptaciones de una especie particular de murciélago pueden influir directamente en los tipos de presas que están disponibles para ella. [129]

El vuelo ha permitido a los murciélagos convertirse en uno de los grupos de mamíferos más ampliamente distribuidos. [130] Aparte del Ártico, la Antártida y unas pocas islas oceánicas aisladas, los murciélagos existen en casi todos los hábitats de la Tierra. [131] Las áreas tropicales tienden a tener más especies que las templadas. [132] Diferentes especies seleccionan diferentes hábitats durante diferentes estaciones, que van desde las costas hasta las montañas y los desiertos, pero requieren refugios adecuados. Los refugios de murciélagos se pueden encontrar en huecos, grietas, follaje e incluso estructuras hechas por el hombre, e incluyen "tiendas" que los murciélagos construyen con hojas. [133] Los megamurciélagos generalmente se posan en los árboles. [134] La mayoría de los micromurciélagos son nocturnos [135] y los megamurciélagos son típicamente diurnos o crepusculares . [136] [137] Se sabe que los microquirópteros exhiben un comportamiento diurno en regiones templadas durante el verano cuando no hay suficiente tiempo nocturno para alimentarse, [138] [139] y en áreas donde hay pocos depredadores aviares durante el día. [140] [141]
En las zonas templadas, algunos microquirópteros migran cientos de kilómetros hasta sus guaridas de hibernación invernal; [142] otros entran en letargo cuando hace frío, despertándose y alimentándose cuando el clima cálido permite que los insectos estén activos. [143] Otros se retiran a cuevas para pasar el invierno e hibernan hasta seis meses. [143] Los microquirópteros rara vez vuelan bajo la lluvia; interfiere con su ecolocalización y no pueden cazar. [144]
Las distintas especies de murciélagos tienen dietas diferentes, que incluyen insectos, néctar, polen, frutas e incluso vertebrados. [145] Los megamurciélagos se alimentan principalmente de frutas, néctar y polen. [136] Debido a su pequeño tamaño, su alto metabolismo y la rápida quema de energía a través del vuelo, los murciélagos deben consumir grandes cantidades de comida para su tamaño. Los murciélagos insectívoros pueden comer más del 120 por ciento de su peso corporal por día, mientras que los murciélagos frugívoros pueden comer más del doble de su peso. [146] Pueden viajar distancias significativas cada noche, excepcionalmente hasta 38,5 km (24 mi) en el caso del murciélago moteado ( Euderma maculatum ), en busca de alimento. [147] Los murciélagos utilizan una variedad de estrategias de caza. [128] Los murciélagos obtienen la mayor parte de su agua de los alimentos que comen; muchas especies también beben de fuentes de agua como lagos y arroyos, volando sobre la superficie y sumergiendo sus lenguas en el agua. [148]
Los quirópteros en su conjunto están en proceso de perder la capacidad de sintetizar vitamina C. [ 149] En una prueba de 34 especies de murciélagos de seis familias principales, incluidas las principales familias de murciélagos que se alimentan de insectos y frutas, se encontró que todas habían perdido la capacidad de sintetizarla, y esta pérdida puede derivar de un ancestro común de los murciélagos, como una única mutación. [150] [b] Al menos dos especies de murciélago, el murciélago frugívoro ( Rousettus leschenaultii ) y el murciélago insectívoro ( Hipposideros armiger ), han conservado su capacidad de producir vitamina C. [151]
La mayoría de los microquirópteros, especialmente en áreas templadas, se alimentan de insectos. [145] La dieta de un murciélago insectívoro puede abarcar muchas especies, [152] incluyendo moscas , mosquitos , escarabajos , polillas, saltamontes , grillos , termitas , abejas , avispas , efímeras y tricópteros . [47] [153] [154] Un gran número de murciélagos mexicanos de cola libre ( Tadarida brasiliensis ) vuelan cientos de metros sobre el suelo en el centro de Texas para alimentarse de polillas migratorias. [155] Las especies que cazan insectos en vuelo, como el pequeño murciélago café ( Myotis lucifugus ), pueden atrapar un insecto en el aire con la boca y comérselo en el aire o usar las membranas de su cola o sus alas para recoger el insecto y llevarlo a la boca. [156] [157] El murciélago también puede llevar al insecto a su refugio y comérselo allí. [158] Las especies de murciélagos de movimiento más lento, como el murciélago orejudo pardo ( Plecotus auritus ) y muchas especies de murciélagos de herradura, pueden tomar o recolectar insectos de la vegetación o cazarlos desde perchas. [47] Los murciélagos insectívoros que viven en latitudes altas tienen que consumir presas con mayor valor energético que los murciélagos tropicales. [159]

La alimentación frugívora, o frugivoría, se encuentra en ambos subórdenes principales. Los murciélagos prefieren la fruta madura, arrancándola de los árboles con los dientes. Vuelan de regreso a sus refugios para comer la fruta, chupando el jugo y escupiendo las semillas y la pulpa en el suelo. Esto ayuda a dispersar las semillas de estos árboles frutales, que pueden echar raíces y crecer donde los murciélagos las han dejado, y muchas especies de plantas dependen de los murciélagos para la dispersión de semillas . [160] [161] Se ha registrado que el murciélago frugívoro jamaiquino ( Artibeus jamaicensis ) lleva frutas que pesan entre 3 y 14 g ( 1 ⁄ 8 – 1 ⁄ 2 oz) o incluso hasta 50 g ( 1+3 ⁄ 4 onzas). [162]
Los murciélagos nectarívoros han adquirido adaptaciones especializadas. Estos murciélagos poseen hocicos largos y lenguas largas y extensibles cubiertas de cerdas finas que les ayudan a alimentarse de determinadas flores y plantas. [161] [163] El murciélago nectarívoro de labios tubulares ( Anoura fistulata ) tiene la lengua más larga de todos los mamíferos en relación con su tamaño corporal. Esto les resulta beneficioso en términos de polinización y alimentación. Sus lenguas largas y estrechas pueden llegar hasta lo profundo de la forma de copa alargada de algunas flores. Cuando la lengua se retrae, se enrolla dentro de la caja torácica. [163] Debido a estas características, los murciélagos nectarívoros no pueden recurrir fácilmente a otras fuentes de alimento en épocas de escasez, lo que los hace más propensos a la extinción que otros tipos de murciélagos. [164] [165] La alimentación con néctar también ayuda a una variedad de plantas, ya que estos murciélagos sirven como polinizadores , ya que el polen se adhiere a su pelaje mientras se alimentan. Alrededor de 500 especies de plantas con flores dependen de la polinización de los murciélagos y, por lo tanto, tienden a abrir sus flores durante la noche. [161] Muchas plantas de la selva tropical dependen de la polinización de los murciélagos. [166]

Algunos murciélagos cazan otros vertebrados, como peces, ranas, lagartijas, aves y mamíferos. [47] [168] El murciélago de labios con flecos ( Trachops cirrhosus, ), por ejemplo, es experto en atrapar ranas. Estos murciélagos localizan grandes grupos de ranas siguiendo sus llamadas de apareamiento, y luego las arrancan de la superficie del agua con sus afilados dientes caninos. [169] El murciélago nóctulo mayor puede atrapar pájaros en vuelo. [167] Algunas especies, como el murciélago bulldog mayor ( Noctilio leporinus ) cazan peces. Utilizan la ecolocalización para detectar pequeñas ondulaciones en la superficie del agua, se lanzan en picado y utilizan garras especialmente agrandadas en sus patas traseras para atrapar el pez, luego llevan a su presa a un refugio de alimentación y la consumen. [170] Se sabe que al menos dos especies de murciélagos se alimentan de otros murciélagos: el murciélago espectral ( Vampyrum spectrum ) y el murciélago fantasma ( Macroderma gigas ). [171]
.jpg/1280px-Desmo-boden_(cropped).jpg)
Unas pocas especies, específicamente los murciélagos vampiros común, de alas blancas y de patas peludas , se alimentan solo de sangre animal ( hematofagia ). El murciélago vampiro común generalmente se alimenta de grandes mamíferos como el ganado ; los vampiros de patas peludas y de alas blancas se alimentan de aves. [172] Los murciélagos vampiros apuntan a presas dormidas y pueden detectar la respiración profunda. [173] Los sensores de calor en la nariz los ayudan a detectar vasos sanguíneos cerca de la superficie de la piel. [174] Perforan la piel del animal con sus dientes, mordiendo un pequeño colgajo, [175] y lamen la sangre con sus lenguas, que tienen surcos laterales adaptados a este propósito. [176] La sangre se evita que se coagule mediante un anticoagulante en la saliva. [175]
Los murciélagos están sujetos a la depredación de aves rapaces , como búhos , halcones y halcones , y en los refugios de depredadores terrestres capaces de trepar, como los gatos. [177] Los murciélagos que vuelan bajo son vulnerables a los cocodrilos . [178] Se sabe que veinte especies de serpientes tropicales del Nuevo Mundo capturan murciélagos, a menudo esperando en las entradas de refugios, como cuevas, a que los murciélagos pasen volando. [179] J. Rydell y JR Speakman argumentan que los murciélagos desarrollaron la nocturnidad durante el período Eoceno temprano y medio para evitar a los depredadores. [177] Algunos zoólogos creen que la evidencia es equívoca hasta ahora. [180]
.jpg/1280px-Little_Brown_Bat_with_White_Nose_Syndrome_(Greeley_Mine,_cropped).jpg)
Como la mayoría de los mamíferos, los murciélagos son huéspedes de una serie de parásitos internos y externos. [181] Entre los ectoparásitos , los murciélagos son portadores de pulgas y ácaros , así como parásitos específicos como chinches y moscas de murciélago ( Nycteribiidae y Streblidae ). [182] [183] Los murciélagos se encuentran entre los pocos órdenes de mamíferos no acuáticos que no albergan piojos , posiblemente debido a la competencia de parásitos más especializados que ocupan el mismo nicho. [183]
El síndrome de la nariz blanca es una afección asociada con la muerte de millones de murciélagos en el este de los Estados Unidos y Canadá. [184] La enfermedad recibe su nombre de un hongo blanco , Pseudogymnoascus destructans , que crece en los hocicos, las orejas y las alas de los murciélagos afectados. El hongo se transmite principalmente de murciélago a murciélago y causa la enfermedad. [185] El hongo se descubrió por primera vez en el centro del estado de Nueva York en 2006 y se extendió rápidamente a todo el este de los EE. UU. al norte de Florida; se han observado tasas de mortalidad del 90 al 100 % en la mayoría de las cuevas afectadas. [186] Nueva Inglaterra y los estados del Atlántico medio han sido testigos, desde 2006, de la extirpación total de especies enteras y otras con números que han pasado de cientos de miles, incluso millones, a unos pocos cientos o menos. [187] Nueva Escocia, Quebec, Ontario y Nuevo Brunswick han sido testigos de muertes idénticas, y el gobierno canadiense ha hecho preparativos para proteger todas las poblaciones de murciélagos restantes en su territorio. [188] La evidencia científica sugiere que los inviernos más largos, donde el hongo tiene un período más largo para infectar a los murciélagos, resultan en una mayor mortalidad. [189] [190] [191] En 2014, la infección cruzó el río Mississippi, [192] y en 2017, se encontró en murciélagos en Texas. [193]
Los murciélagos son reservorios naturales de una gran cantidad de patógenos zoonóticos , [194] incluyendo la rabia , endémica en muchas poblaciones de murciélagos, [195] [196] [197] la histoplasmosis tanto directamente como en el guano, [198] los virus Nipah y Hendra , [199] [200] y posiblemente el virus del ébola , [201] [202] cuyo reservorio natural aún se desconoce. [203] [204] Su alta movilidad, amplia distribución, larga vida, simpatría sustancial (superposición de rangos) de especies y comportamiento social hacen que los murciélagos sean huéspedes favorables y vectores de enfermedades. [205] Las revisiones han encontrado diferentes respuestas en cuanto a si los murciélagos tienen más virus zoonóticos que otros grupos de mamíferos. Una revisión de 2015 concluyó que los murciélagos, roedores y primates albergaban significativamente más virus zoonóticos (que pueden transmitirse a los humanos) que otros grupos de mamíferos, aunque las diferencias entre los tres grupos mencionados no fueron significativas (los murciélagos no tienen más virus zoonóticos que los roedores y los primates). [206] Otra revisión de 2020 sobre mamíferos y aves concluyó que la identidad de los grupos taxonómicos no tenía ningún impacto en la probabilidad de albergar virus zoonóticos. En cambio, los grupos más diversos tenían una mayor diversidad viral. [207]
Parecen ser muy resistentes a muchos de los patógenos que portan, lo que sugiere un grado de adaptación a sus sistemas inmunológicos. [205] [208] [209] Sus interacciones con el ganado y las mascotas, incluida la depredación por murciélagos vampiros, los encuentros accidentales y la carroña de cadáveres de murciélagos, agravan el riesgo de transmisión zoonótica. [196] Los murciélagos están implicados en la aparición del síndrome respiratorio agudo severo (SARS) en China, ya que sirven como huéspedes naturales de los coronavirus , varios de una sola cueva en Yunnan , uno de los cuales se convirtió en el virus del SARS. [198] [210] [211] Sin embargo, no causan ni propagan COVID-19 . [212]
Algunos murciélagos llevan vidas solitarias, mientras que otros viven en colonias de más de un millón. [213] Por ejemplo, el murciélago mexicano de cola libre vuela más de mil millas hasta la cueva de 100 pies (30 m) de ancho conocida como Bracken Cave cada marzo a octubre, que alberga a unos asombrosos veinte millones de la especie, [214] mientras que un murciélago ratonero vive una vida casi completamente solitaria. [215] Vivir en grandes colonias reduce el riesgo de depredación para un individuo. [47] Las especies de murciélagos templados pueden enjambrar en los sitios de hibernación a medida que se acerca el otoño. Esto puede servir para introducir a las crías en los sitios de hibernación, señalar la reproducción en los adultos y permitir que los adultos se reproduzcan con los de otros grupos. [216]
Varias especies tienen una estructura social de fisión-fusión , donde un gran número de murciélagos se congregan en una zona de descanso, junto con la separación y mezcla de subgrupos. Dentro de estas sociedades, los murciélagos pueden mantener relaciones a largo plazo. [217] Algunas de estas relaciones consisten en hembras relacionadas matrilinealmente y sus crías dependientes. [218] El intercambio de alimentos y el acicalamiento mutuo pueden ocurrir en ciertas especies, como el murciélago vampiro común ( Desmodus rotundus ), y estos fortalecen los lazos sociales. [219] [220] Se ha observado felación homosexual en el zorro volador de Bonin Pteropus pselaphon [221] y el zorro volador indio Pteropus medius , [222] aunque la función y el propósito de este comportamiento no están claros.

Los murciélagos se encuentran entre los mamíferos más vocales y producen llamadas para atraer parejas, encontrar compañeros de descanso y defender recursos. Estas llamadas son típicamente de baja frecuencia y pueden viajar largas distancias. [47] [224] Los murciélagos de cola libre mexicanos son una de las pocas especies que "cantan" como los pájaros. Los machos cantan para atraer a las hembras. Las canciones tienen tres frases: chirridos, trinos y zumbidos, los primeros tienen sílabas "A" y "B". Las canciones de los murciélagos son altamente estereotipadas pero con variación en el número de sílabas, el orden de las frases y las repeticiones de frases entre individuos. [223] Entre los murciélagos de nariz de lanza mayor ( Phyllostomus hastatus ), las hembras producen llamadas fuertes y de banda ancha entre sus compañeros de descanso para formar la cohesión del grupo. Las llamadas difieren entre los grupos de descanso y pueden surgir del aprendizaje vocal. [225]
En un estudio sobre murciélagos frugívoros egipcios cautivos, los investigadores pudieron identificar el 70% de los llamados dirigidos en cuanto a qué murciélago individual los emitió, y el 60% se pudo categorizar en cuatro contextos: peleas por comida, empujones por la posición en su grupo de sueño, protestas por intentos de apareamiento y discusiones cuando estaban posados muy cerca uno del otro. Los animales emitían sonidos ligeramente diferentes cuando se comunicaban con diferentes murciélagos individuales, especialmente aquellos del sexo opuesto. [226] En el murciélago martillo altamente dimórfico sexualmente ( Hypsignathus monstrosus ), los machos producen llamados profundos, resonantes y monótonos para atraer a las hembras. Los murciélagos en vuelo emiten señales vocales para el control del tráfico. Los murciélagos bulldog mayor graznan cuando están en curso de colisión entre sí. [224]
Los murciélagos también se comunican por otros medios. Los murciélagos machos de hombros amarillos pequeños ( Sturnira lilium ) tienen glándulas en los hombros que producen un olor picante durante la temporada de reproducción. Como muchas otras especies, tienen pelos especializados para retener y dispersar secreciones. Dichos pelos forman un llamativo collar alrededor del cuello de algunos murciélagos megamachos del Viejo Mundo. Los murciélagos machos de alas en saco mayores ( Saccopteryx bilineata ) tienen sacos en sus alas en los que mezclan secreciones corporales como saliva y orina para crear un perfume que esparcen en los sitios de descanso, un comportamiento conocido como "salado". El salado puede ir acompañado de cantos. [224]

La mayoría de las especies de murciélagos son polígamas , es decir , los machos se aparean con varias hembras . Los murciélagos ...
En los murciélagos que viven en zonas templadas, el apareamiento se produce a finales de verano y principios de otoño. [233] Los murciélagos tropicales pueden aparearse durante la estación seca. [234] Después de la cópula, el macho puede dejar un tapón de apareamiento para bloquear el esperma de otros machos y asegurar así su paternidad. [235] En las especies que hibernan, se sabe que los machos se aparean con las hembras en letargo. [47] Las hembras de murciélago utilizan una variedad de estrategias para controlar el momento del embarazo y el nacimiento de las crías, para que el parto coincida con la máxima capacidad alimentaria y otros factores ecológicos. Las hembras de algunas especies tienen una fertilización retardada, en la que el esperma se almacena en el tracto reproductivo durante varios meses después del apareamiento. El apareamiento ocurre a finales de verano y principios de otoño, pero la fertilización no ocurre hasta finales de invierno y principios de primavera siguientes. Otras especies presentan una implantación retardada , en la que el óvulo se fertiliza después del apareamiento, pero permanece libre en el tracto reproductivo hasta que las condiciones externas se vuelven favorables para dar a luz y cuidar a la descendencia. [236] En otra estrategia, se produce la fecundación y la implantación, pero el desarrollo del feto se retrasa hasta que se dan las condiciones adecuadas. Durante el retraso del desarrollo, la madre mantiene vivo el óvulo fecundado con nutrientes. Este proceso puede continuar durante un largo período, gracias al avanzado sistema de intercambio de gases. [237]

En el caso de los murciélagos que viven en zonas templadas, los nacimientos suelen tener lugar en mayo o junio en el hemisferio norte; en el hemisferio sur, en noviembre y diciembre. Las especies tropicales dan a luz al comienzo de la temporada de lluvias. [238] En la mayoría de las especies de murciélagos, las hembras llevan y dan a luz una sola cría por camada. [239] Al nacer, una cría de murciélago puede pesar hasta el 40 por ciento del peso de la madre, [47] y la cintura pélvica de la hembra puede expandirse durante el parto, ya que las dos mitades están conectadas por un ligamento flexible. [240] Las hembras suelen dar a luz con la cabeza hacia arriba o en posición horizontal, utilizando la gravedad para facilitar el parto. La cría emerge primero de atrás, posiblemente para evitar que las alas se enreden, y la hembra la acuna en las membranas de sus alas y cola. En muchas especies, las hembras dan a luz y crían a sus crías en colonias de maternidad y pueden ayudarse entre sí en el parto. [241] [242] [240]
La mayor parte del cuidado de un murciélago joven proviene de la madre. En las especies monógamas, el padre desempeña un papel. La aloamamantamiento, donde una hembra amamanta a la cría de otra madre, ocurre en varias especies. Esto puede servir para aumentar el tamaño de la colonia en especies donde las hembras regresan a su colonia natal para reproducirse. [47] La capacidad de un murciélago joven para volar coincide con el desarrollo de un cuerpo adulto y la longitud de las extremidades anteriores. Para el murciélago marrón pequeño, esto ocurre alrededor de dieciocho días después del nacimiento. El destete de las crías en la mayoría de las especies ocurre en menos de ochenta días. El murciélago vampiro común amamanta a sus crías más allá de eso y los murciélagos vampiro jóvenes alcanzan la independencia más tarde en la vida que otras especies. Esto probablemente se debe a la dieta basada en sangre de la especie, que es difícil de obtener todas las noches. [243]

La esperanza de vida máxima de los murciélagos es tres veces y media mayor que la de otros mamíferos de tamaño similar. Se han registrado seis especies que viven más de treinta años en estado salvaje: el murciélago orejudo pardo ( Plecotus auritus ), el murciélago pardo chico ( Myotis lucifugus ), el murciélago siberiano ( Myotis sibiricus ), el murciélago ratonero menor ( Myotis blythii ), el murciélago de herradura mayor ( Rhinolophus ferrumequinum ) y el zorro volador indio ( Pteropus giganteus ). [244] Una hipótesis consistente con la teoría de la tasa de vida vincula esto con el hecho de que reducen su tasa metabólica mientras hibernan ; los murciélagos que hibernan, en promedio, tienen una esperanza de vida más larga que los murciélagos que no lo hacen. [245] [246]
Otra hipótesis es que volar ha reducido su tasa de mortalidad, lo que también sería cierto para las aves y los mamíferos planeadores. Las especies de murciélagos que dan a luz a múltiples crías generalmente tienen una vida más corta que las especies que dan a luz a una sola cría. Las especies que se posan en cuevas pueden tener una vida más larga que las especies que no lo hacen debido a la menor depredación en cuevas. Un murciélago siberiano macho fue recapturado en estado salvaje después de 41 años, lo que lo convierte en el murciélago más antiguo conocido. [246] [247]
Estados de conservación de los murciélagos a partir de 2020 según la UICN (1.314 especies en total) [248]
Groups such as the Bat Conservation International[249] aim to increase awareness of bats' ecological roles and the environmental threats they face. In the United Kingdom, all bats are protected under the Wildlife and Countryside Acts, and disturbing a bat or its roost can be punished with a heavy fine.[250]In Sarawak, Malaysia, "all bats"[251] are protected under the Wildlife Protection Ordinance 1998,[251] but species such as the hairless bat (Cheiromeles torquatus) are still eaten by the local communities.[252] Humans have caused the extinction of several species of bat in modern history, the most recent being the Christmas Island pipistrelle (Pipistrellus murrayi), which was declared extinct in 2009.[253]
Many people put up bat houses to attract bats.[254] The 1991 University of Florida bat house is the largest occupied artificial roost in the world, with around 400,000 residents.[255] In Britain, thickwalled and partly underground World War II pillboxes have been converted to make roosts for bats,[256][257] and purpose-built bat houses are occasionally built to mitigate damage to habitat from road or other developments.[258][259] Cave gates are sometimes installed to limit human entry into caves with sensitive or endangered bat species. The gates are designed not to limit the airflow, and thus to maintain the cave's micro-ecosystem.[260] Of the 47 species of bats found in the United States, 35 are known to use human structures, including buildings and bridges. Fourteen species use bat houses.[261]
Bats are eaten in countries across Africa, Asia and the Pacific Rim. In some cases, such as in Guam, flying foxes have become endangered through being hunted for food.[262] There is evidence that suggests that wind turbines might create sufficient barotrauma (pressure damage) to kill bats.[263] Bats have typical mammalian lungs, which are thought to be more sensitive to sudden air pressure changes than the lungs of birds, making them more liable to fatal rupture.[264][265][266][267][268] Bats may be attracted to turbines, perhaps seeking roosts, increasing the death rate.[264] Acoustic deterrents may help to reduce bat mortality at wind farms.[269]
The diagnosis and contribution of barotrauma to bat deaths near wind turbine blades have been disputed by other research comparing dead bats found near wind turbines with bats killed by impact with buildings in areas with no turbines.[270]
,_from_Los_Caprichos_-_Google_Art_Project.jpg/1280px-Francisco_Jos%C3%A9_de_Goya_y_Lucientes_-_The_sleep_of_reason_produces_monsters_(No._43),_from_Los_Caprichos_-_Google_Art_Project.jpg)
Since bats are mammals, yet can fly, they are considered to be liminal beings in various traditions.[271] In many cultures, including in Europe, bats are associated with darkness, death, witchcraft, and malevolence.[272] Among Native Americans such as the Creek, Cherokee and Apache, the bat is identified as a trickster.[273] In Tanzania, a winged batlike creature known as Popobawa is believed to be a shapeshifting evil spirit that assaults and sodomises its victims.[274] In Aztec mythology, bats symbolised the land of the dead, destruction, and decay.[275][276][277] An East Nigerian tale tells that the bat developed its nocturnal habits after causing the death of his partner, the bush-rat, and now hides by day to avoid arrest.[278]
More positive depictions of bats exist in some cultures. In China, bats have been associated with happiness, joy and good fortune. Five bats are used to symbolise the "Five Blessings": longevity, wealth, health, love of virtue and peaceful death.[279] The bat is sacred in Tonga and is often considered the physical manifestation of a separable soul.[280] In the Zapotec civilisation of Mesoamerica, the bat god presided over corn and fertility.[281]

The Weird Sisters in Shakespeare's Macbeth used the fur of a bat in their brew.[282] In Western culture, the bat is often a symbol of the night and its foreboding nature. The bat is a primary animal associated with fictional characters of the night, both villainous vampires, such as Count Dracula and before him Varney the Vampire,[283] and heroes, such as the DC Comics character Batman.[284] Kenneth Oppel's Silverwing novels narrate the adventures of a young bat,[285] based on the silver-haired bat of North America.[286]
The bat is sometimes used as a heraldic symbol in Spain and France, appearing in the coats of arms of the towns of Valencia, Palma de Mallorca, Fraga, Albacete, and Montchauvet.[287][288][289] Three US states have an official state bat. Texas and Oklahoma are represented by the Mexican free-tailed bat, while Virginia is represented by the Virginia big-eared bat (Corynorhinus townsendii virginianus).[290]
Insectivorous bats in particular are especially helpful to farmers, as they control populations of agricultural pests and reduce the need to use pesticides. It has been estimated that bats save the agricultural industry of the United States anywhere from $3.7 billion to $53 billion per year in pesticides and damage to crops. This also prevents the overuse of pesticides, which can pollute the surrounding environment, and may lead to resistance in future generations of insects.[291]
Bat dung, a type of guano, is rich in nitrates and is mined from caves for use as fertiliser.[292] During the US Civil War, saltpetre was collected from caves to make gunpowder. At the time, it was believed that the nitrate all came from the bat guano, but it is now known that most of it is produced by nitrifying bacteria.[293]
The Congress Avenue Bridge in Austin, Texas, is the summer home to North America's largest urban bat colony, an estimated 1,500,000 Mexican free-tailed bats. About 100,000 tourists a year visit the bridge at twilight to watch the bats leave the roost.[294]
Several molecular studies have shown that Chiroptera belong to the Laurasiatheria (represented by carnivores, pangolins, cetartiodactyls, eulipotyphlans, and perissodactyls) and are only distantly related to dermopterans, scandentians, and primates. (Nikaido et al. 2000; Lin and Penny 2001; Madsen et al. 2001; Murphy et al. 2001a, 2001b; Van Den Bussche and Hoofer 2004)
{{cite journal}}: CS1 maint: DOI inactive as of September 2024 (link)A. fistulata (shown lapping sugar water from a tube) has the longest tongue, relative to body length, of any mammal – and now scientists think they know why.
The currently known viruses that have been found in bats are reviewed and the risks of transmission to humans are highlighted.
Bats, which represent approximately 24% of all known mammalian species, frequently act as vectors of lyssaviruses.
In this paper we describe the isolation of HeV from pteropid bats, corroborating our serological and epidemiological evidence that these animals are a natural reservoir host of this virus.
We find evidence of asymptomatic infection by Ebola virus in three species of megabats, indicating that these animals may be acting as a reservoir for this deadly virus.
Long known as vectors for rabies, bats may be the origin of some of the most deadly emerging viruses, including SARS, Ebola, Nipah, Hendra and Marburg.Note: This is a lay summary of the various scientific publications cited in the preceding sentence.
Scientists do not know where Ebola virus comes from.
Despite concerted investigative efforts, the natural reservoir of the virus is unknown.
The genetic diversity of bat-derived sequences supports the notion that bats are a natural reservoir host of the SARS cluster of coronaviruses.
"Shade is based on a Silver-Haired Bat. I thought they were very dashing-looking creatures. I liked the fact this was a bat that lived in the same part of the world as me (eastern Canada). These are small creatures, with a wing span of a few inches. Their bodies are about the same size as mice. They're insectivores, which means they eat only insects." – K.O.
{{cite book}}: CS1 maint: location missing publisher (link)