Clase de mecanorreceptores que se encuentran en los insectos.
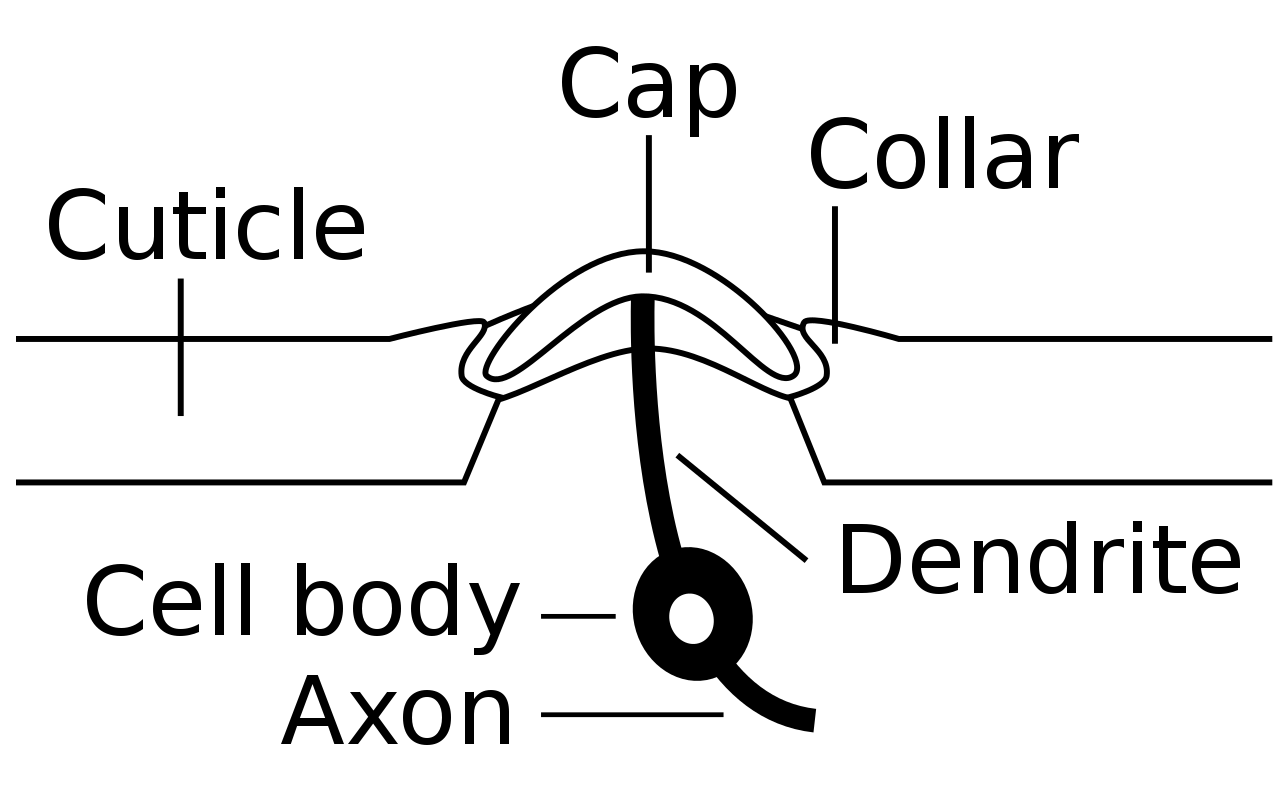
Corte transversal de un sensillum campaniforme. Cada sensillum está incrustado en una cavidad cuticular e inervado por una única neurona sensorial. La neurona se excita cuando la tensión en la cutícula deforma los bordes de la cavidad (collar) y marca la cúpula cuticular flexible (tapa).Panel superior: Micrografía electrónica de barrido (SEM) de sensilla campaniforme en el tarso de Drosophila melanogaster . Panel inferior: MEB de campaniforme en la base del halter de una mosca sarcófaga.
Las sensillas campaniformes son una clase de mecanorreceptores que se encuentran en los insectos y que responden al estrés local y a la tensión dentro de la cutícula del animal. Las sensillas campaniformes funcionan como propioceptores que detectan la carga mecánica como resistencia a la contracción muscular, [1] [2] similar a los órganos tendinosos de Golgi de los mamíferos . [3] [4] La retroalimentación sensorial de la sensilla campaniforme está integrada en el control de la postura y la locomoción. [5] [6]
Estructura
Cada sensillum campaniforme consta de una cúpula flexible, que está incrustada en una cavidad esponjosa dentro de la cutícula e inervada por las dendritas de una única neurona sensorial bipolar (ver sección transversal esquemática). Las sensillas campaniformes suelen tener forma ovalada con ejes largos de aproximadamente 5 a 10 μm (ver SEM).
Las sensilla campaniformes se distribuyen por la superficie del cuerpo de muchos insectos. La mosca de la fruta Drosophila melanogaster , por ejemplo, tiene más de 680 sensillas. [7] Las sensilla campaniformes se encuentran en regiones donde es probable que el estrés sea alto, incluidas las patas, las antenas, las alas y los halterios . [7] [8] [9] La sensilla puede aparecer sola, pero las sensilla con orientaciones similares a menudo se agrupan.
Distribución de grupos de sensillas campaniformes en una pata de insecto palo (vista anterior). El recuadro muestra una vista superior de los dos grupos del trocánter dorsal (G3 y G4). Las sensilla de estos grupos tienen orientaciones mutuamente perpendiculares. Cada sensillum se excita preferentemente por compresión a lo largo de su eje corto (flechas). El grupo proximal (G3) está orientado perpendicularmente al eje longitudinal del trocánter y se excita cuando el trocánter-fémur se dobla hacia arriba. El grupo más distal (G4) está orientado en paralelo al eje longitudinal del trocánter y se excita cuando el trocánter-fémur se dobla hacia abajo.
Sensilla campaniforme en las piernas
En las piernas, los grupos de sensillas campaniformes se encuentran cerca de las articulaciones en todos los segmentos excepto en la coxa (ver esquema de la pierna), y la mayoría de las sensillas se encuentran en el trocánter proximal. [10] El número y la ubicación de las sensillas en las patas varía poco entre individuos de la misma especie, [7] y se pueden encontrar grupos homólogos de sensillas entre especies. [10]
Sensilla campaniforme en alas y halteres
Distribución de sensilla campaniforme (CS) en el ala y halter de Drosophila melanogaster . Adaptado de Aiello et al. (2021). [9]
Las sensilla campaniformes suelen aparecer en ambos lados del ala (ver esquema del ala). El número exacto y la ubicación varían ampliamente entre especies, lo que probablemente refleja diferencias en el comportamiento de vuelo. [9] Sin embargo, entre especies, la mayoría de las sensillas campaniformes se encuentran cerca de la base del ala. [9] Los modelos computacionales predicen que esta es una ubicación óptima para detectar las rotaciones del cuerpo durante el vuelo, siendo el rendimiento de la detección robusto a las perturbaciones externas y la pérdida de sensores. [11]
En dípteros como Drosophila , la mayor densidad de sensillas campaniformes se encuentra en la base de las alas traseras modificadas, los halterios (ver esquema de los halterios). [7] [8]
Función
Propiedades de respuesta
Cuando las deformaciones cuticulares comprimen un sensillum campaniforme, los bordes de la cavidad (cuello) marcan la tapa cuticular. [12] Esto aprieta la punta dendrítica de la neurona sensorial y abre sus canales de mecanotransducción (de la familia TRP [13] ), lo que conduce a la generación de potenciales de acción que se transmiten al cordón nervioso ventral , el insecto análogo al vertebrado. médula espinal.
La actividad de la sensilla campaniforme fue registrada por primera vez por John William Sutton Pringle a finales de la década de 1930. [14] Pringle también determinó que la forma ovalada de muchas sensillas las hace direccionalmente selectivas [15] : responden mejor a la compresión a lo largo de su eje corto. Por lo tanto, incluso las sensillas vecinas pueden tener sensibilidades muy diferentes a la tensión dependiendo de su orientación en la cutícula. Por ejemplo, los insectos palo poseen dos grupos de sensillas campaniformes en el lado dorsal del trocánter de sus patas cuyos ejes cortos están orientados perpendicularmente entre sí [1] (ver recuadro en el esquema de las patas). Como resultado, un grupo (G3) responde cuando la pierna se dobla hacia arriba, mientras que el otro grupo (G4) responde cuando la pierna se dobla hacia abajo. La sensilla campaniforme redonda puede ser sensible en todas las direcciones [16] o mostrar sensibilidad direccional si la tapa está acoplada asimétricamente con el collar circundante. [17]
La actividad de la sensilla campaniforme puede ser de adaptación lenta (tónica), lo que indica la magnitud de la deformación cuticular, y/o de adaptación rápida (fásica), lo que indica la velocidad de la deformación cuticular. [1] [18] Según sus respuestas a los estímulos de ruido blanco, la sensilla campaniforme también puede describirse de manera más general como la señalización de dos características que se aproximan a la derivada entre sí. [19] Esto sugiere que las propiedades de respuesta neuronal de las sensillas son bastante genéricas y que la especialización funcional surge principalmente de cómo las sensillas están incrustadas en la cutícula. [19] [20] Además, la actividad se adapta a cargas constantes y muestra histéresis (dependencia histórica) en respuesta a la carga cíclica. [18]
Las sensillas campaniformes se proyectan directamente a las neuronas motoras [21] y a varias interneuronas, que integran sus señales con señales de otros propioceptores. [22] De esta manera, la actividad de la sensilla campaniforme puede afectar la magnitud y el momento de las contracciones musculares. [5]
Función de la sensilla campaniforme de la pierna.
Las sensillas campaniformes de las piernas se activan al estar de pie y al caminar. [23] [24] Se cree que su retroalimentación sensorial refuerza la actividad muscular durante la fase de postura [1] [24] [25] y contribuye a la coordinación entre las piernas, [26] [27] muy similar a la retroalimentación sensorial del aparato de Golgi de los mamíferos. órganos tendinosos . [28] [29] La retroalimentación de la sensilla campaniforme de la pierna también es importante para el control de las patadas y los saltos. [30] [31]
Función del ala y del halter campaniforme sensilla
Las sensillas campaniformes en las alas y los halterios se activan a medida que estas estructuras oscilan hacia adelante y hacia atrás durante el vuelo, y la fase de activación depende de la ubicación de las sensillas. [9] [32] La sensilla campaniforme del ala codifica las fuerzas aerodinámicas e inerciales del ala, mientras que se cree que la sensilla de la base del halter codifica las fuerzas de Coriolis inducidas por la rotación del cuerpo durante el vuelo, lo que permite que la estructura funcione como un giroscopio . [33] Se cree que la retroalimentación de la sensilla campaniforme del ala y del halter media los reflejos compensatorios para mantener el equilibrio durante el vuelo. [34] [35]
Modelos computacionales
Para comprender mejor la función de la sensilla campaniforme, se están desarrollando modelos computacionales que imitan sus propiedades de respuesta para su uso en simulaciones y robótica. [36] [37] En patas robóticas, los modelos pueden filtrar la entrada de sensores de tensión diseñados "estilo campaniforme sensilla" en tiempo real. [38] Una ventaja de este filtrado bioinspirado es que permite la adaptación a la carga con el tiempo (ver arriba), lo que hace que los sensores de tensión esencialmente se autocalibran a diferentes cargas transportadas por el robot. [38]
Referencias
^ abcd Zill SN, Schmitz J, Chaudhry S, Büschges A (septiembre de 2012). "La codificación de fuerza en las patas de los insectos palo delinea un marco de referencia para el control motor". Revista de Neurofisiología . 108 (5): 1453–72. doi :10.1152/jn.00274.2012. PMC 3774582 . PMID 22673329.
^ Zill SN, Chaudhry S, Büschges A, Schmitz J (noviembre de 2013). "Especificidad direccional y codificación de fuerzas y cargas musculares por sensilla campaniforme tibial del insecto palo, incluidos receptores con tapas cuticulares redondas". Estructura y desarrollo de artrópodos . 42 (6): 455–467. doi :10.1016/j.asd.2013.10.001. PMID 24126203.
^ Duysens J, Clarac F, Cruse H (enero de 2000). "Mecanismos de regulación de carga en la marcha y la postura: aspectos comparativos". Revisiones fisiológicas . 80 (1): 83-133. doi :10.1152/physrev.2000.80.1.83. PMID 10617766.
^ Tuthill JC, Azim E (marzo de 2018). "Propiocepción". Biología actual . 28 (5): R194–R203. doi : 10.1016/j.cub.2018.01.064 . PMID 29510103.
^ ab Zill S, Schmitz J, Büschges A (julio de 2004). "Sensación de carga y control de postura y locomoción". Estructura y desarrollo de artrópodos . 33 (3): 273–86. doi :10.1016/j.asd.2004.05.005. PMID 18089039.
^ Tuthill JC, Wilson RI (octubre de 2016). "Mecanosensación y control motor adaptativo en insectos". Biología actual . 26 (20): R1022–R1038. doi :10.1016/j.cub.2016.06.070. PMC 5120761 . PMID 27780045.
^ abcd Dinges GF, Chockley AS, Bockemühl T, Ito K, Blanke A, Büschges A (julio de 2020). "Ubicación y disposición de sensilla campaniforme en Drosophila melanogaster". La Revista de Neurología Comparada . 529 (4): 905–925. doi : 10.1002/cne.24987 . PMID 32678470.
^ ab Agrawal, Sweta; Grimaldi, David; Zorro, Jessica L. (1 de marzo de 2017). "Morfología de haltere y disposición campaniforme de sensilla en Diptera". Estructura y desarrollo de artrópodos . 46 (2): 215–229. doi : 10.1016/j.asd.2017.01.005 . ISSN 1467-8039. PMID 28161605.
^ abcde Aiello, Brett R.; Stanchak, Kathryn E.; Weber, Alison I.; Deora, Tanvi; Sponberg, Simón; Brunton, Bingni W. (24 de junio de 2021). "Distribución espacial de mecanosensores sensilla campaniformes en las alas: forma, función y filogenia". Opinión actual en ciencia de insectos . 48 : 8-17. doi : 10.1016/j.cois.2021.06.002 . ISSN 2214-5745. PMID 34175464.
^ ab Harris CM, Dinges GF, Haberkorn A, Gebehart C, Büschges A, Zill SN (septiembre de 2020). "Gradientes en mecanotransducción de fuerza y peso corporal en insectos". Estructura y desarrollo de artrópodos . 58 : 100970. doi : 10.1016/j.asd.2020.100970 . PMID 32702647.
^ Weber, Alison I.; Daniel, Thomas L.; Brunton, Bingni W. (11 de agosto de 2021). Webb, Bárbara (ed.). "La estructura del ala y la codificación neuronal determinan conjuntamente las estrategias de detección en el vuelo de los insectos". PLOS Biología Computacional . 17 (8): e1009195. Código Bib : 2021PLSCB..17E9195W. doi : 10.1371/journal.pcbi.1009195 . ISSN 1553-7358. PMC 8382179 . PMID 34379622.
^ Spinola SM, Chapman KM (1 de septiembre de 1975). "Sangría propioceptiva de la sensilla campaniforme de patas de cucaracha". Revista de fisiología comparada . 96 (3): 257–272. doi :10.1007/BF00612698. ISSN 1432-1351. S2CID 8017950.
^ Liang X, Madrid J, Saleh HS, Howard J (enero de 2011). "NOMPC, un miembro de la familia de canales TRP, se localiza en el cuerpo tubular y el cilio distal de las células receptoras cordotonales y campaniformes de Drosophila". Citoesqueleto . 68 (1): 1–7. doi :10.1002/cm.20493. PMC 3048163 . PMID 21069788.
^ Pringle JW (1938). "Propiocepción en insectos I. Un nuevo tipo de receptor mecánico de los palpos de la cucaracha". Revista de biología experimental . 15 (1): 101–113. doi : 10.1242/jeb.15.1.101 .
^ Pringle JW (1938). "Propiocepción en insectos II. La acción de la sensilla campaniforme en las patas". Revista de biología experimental . 15 (1): 114-131. doi : 10.1242/jeb.15.1.114 .
^ Dickinson MH (1 de agosto de 1992). "Sensibilidad direccional y dinámica de acoplamiento mecánico de la sensilla campaniforme durante las deformaciones en sentido de las cuerdas del ala de la mosca". Revista de biología experimental . 169 (1): 221–233. doi :10.1242/jeb.169.1.221. ISSN 0022-0949.
^ Zill SN, Büschges A, Schmitz J (agosto de 2011). "La codificación de la fuerza aumenta y disminuye por la sensilla campaniforme tibial en el insecto palo, Carausius morosus". Revista de fisiología comparada A: neuroetología, fisiología sensorial, neuronal y conductual . 197 (8): 851–67. doi :10.1007/s00359-011-0647-4. PMID 21544617. S2CID 20865515.
^ ab Ridgel AL, Frazier SF, DiCaprio RA, Zill SN (abril de 2000). "Codificación de fuerzas por sensilla campaniforme tibial de cucaracha: implicaciones en el control dinámico de la postura y la locomoción". Revista de fisiología comparada A: fisiología sensorial, neuronal y conductual . 186 (4): 359–74. doi :10.1007/s003590050436. PMID 10798724. S2CID 12306005.
^ ab Dickerson, Bradley H.; Fox, Jessica L.; Sponberg, Simon (28 de noviembre de 2020). "Diversidad funcional de la codificación genérica en insectos campaniformes sensilla". Opinión actual en fisiología . 19 : 194-203. doi : 10.1016/j.cophys.2020.11.004 . ISSN 2468-8673.
^ Dinges, Gesa F.; Bockemühl, Till; Iacoviello, Francesco; Esquila, Paul R.; Büschges, Ansgar; Blanke, Alexander (mayo de 2022). "La biomecánica de ultra alta resolución sugiere que las subestructuras dentro de los mecanosensores de insectos afectan decisivamente su sensibilidad". Revista de la interfaz de la Royal Society . 19 (190): 20220102. doi :10.1098/rsif.2022.0102. ISSN 1742-5662. PMC 9065962 . PMID 35506211.
^ Phelps, Jasper S.; Hildebrand, David Grant Colburn; Graham, Brett J.; Kuan, Aaron T.; Thomas, Logan A.; Nguyen, Tri M.; Buhmann, Julia; Azevedo, Antonio W.; Sustar, Ana; Agrawal, Sweta; Liu, Mingguan; Shanny, Brendan L.; Funke, enero; Tuthill, John C.; Lee, Wei-Chung Allen (4 de febrero de 2021). "Reconstrucción de circuitos de control motor en Drosophila adulta mediante microscopía electrónica de transmisión automatizada". Celúla . 184 (3): 759–774.e18. doi :10.1016/j.cell.2020.12.013. ISSN 0092-8674. PMC 8312698 . PMID 33400916.
^ Gebehart, Corinna; Schmidt, Joaquín; Büschges, Ansgar (1 de mayo de 2021). "Procesamiento distribuido de retroalimentación de carga y movimiento en la red premotora que controla la articulación de una pata de insecto". Revista de Neurofisiología . 125 (5): 1800–1813. doi :10.1152/jn.00090.2021. ISSN 0022-3077. PMID 33788591. S2CID 232480916.
^ Keller, Bridget R.; Duque, Elizabeth R.; Amer, Aymán S.; Zill, Sasha N. (agosto de 2007). "Ajustar la postura a la carga corporal: las disminuciones de la carga producen señales sensoriales discretas en las patas de las cucarachas que se encuentran libremente". Revista de fisiología comparada A: neuroetología, fisiología sensorial, neuronal y conductual . 193 (8): 881–891. doi :10.1007/s00359-007-0241-y. ISSN 0340-7594. PMID 17541783. S2CID 44044892.
^ ab Pearson KG (1972). "Programación central y control de reflejos del caminar en la cucaracha". Revista de biología experimental . 56 (1): 173–193. doi :10.1242/jeb.56.1.173. ISSN 0022-0949.
^ Zill SN, Chaudhry S, Büschges A, Schmitz J (noviembre de 2015). "La retroalimentación de fuerza refuerza las sinergias musculares en las patas de los insectos". Estructura y desarrollo de artrópodos . 44 (6 puntos A): 541–53. doi :10.1016/j.asd.2015.07.001. PMID 26193626.
^ Zill SN, Keller BR, Duke ER (mayo de 2009). "Las señales sensoriales de descarga en una pierna siguen al inicio de la postura en la otra pierna: transferencia de carga y coordinación emergente al caminar como cucaracha". Revista de Neurofisiología . 101 (5): 2297–304. doi :10.1152/jn.00056.2009. PMID 19261716. S2CID 14691776.
^ Dallmann CJ, Hoinville T, Dürr V, Schmitz J (diciembre de 2017). "Un mecanismo basado en carga para la coordinación entre patas en insectos". Actas. Ciencias Biologicas . 284 (1868): 20171755. doi :10.1098/rspb.2017.1755. PMC 5740276 . PMID 29187626.
^ Pearson KG (enero de 2008). "Papel de la retroalimentación sensorial en el control de la duración de la postura en gatos que caminan". Reseñas de investigaciones sobre el cerebro . Redes en movimiento. 57 (1): 222–7. doi : 10.1016/j.brainresrev.2007.06.014. PMID 17761295. S2CID 23068577.
^ Ekeberg O, Pearson K (diciembre de 2005). "Simulación por computadora de pisar las patas traseras del gato: un examen de los mecanismos que regulan la transición de la postura al balanceo". Revista de Neurofisiología . 94 (6): 4256–68. doi :10.1152/jn.00065.2005. PMID 16049149. S2CID 7185159.
^ Burrows M, Pflüger HJ (1 de julio de 1988). "Bucles de retroalimentación positiva de los propioceptores implicados en los movimientos de las patas de la langosta". Revista de fisiología comparada A. 163 (4): 425–440. doi :10.1007/BF00604897. S2CID 25848693.
^ Norman AP (agosto de 1996). "Retroalimentación propioceptiva en las patadas y saltos de las langostas durante la maduración". Revista de fisiología comparada A: fisiología sensorial, neuronal y conductual . 179 (2): 195-205. doi :10.1007/BF00222786. PMID 8765558. S2CID 2312224.
^ DICKINSON, MICHAEL H. (1 de julio de 1990). "Comparación de las propiedades de codificación de Campaniform Sensilla en el ala de la mosca". Revista de biología experimental . 151 (1): 245–261. doi :10.1242/jeb.151.1.245. ISSN 0022-0949.
^ Pringle, John William Sutton; Gray, James (2 de noviembre de 1948). "El mecanismo giroscópico de los halterios de Diptera". Transacciones filosóficas de la Royal Society de Londres. Serie B, Ciencias Biológicas . 233 (602): 347–384. Código bibliográfico : 1948RSPTB.233..347P. doi : 10.1098/rstb.1948.0007 .
^ Dickinson MH (mayo de 1999). van Leeuwen JL (ed.). "Reflejos de equilibrio mediados por halterio de la mosca de la fruta, Drosophila melanogaster". Transacciones filosóficas de la Royal Society de Londres. Serie B, Ciencias Biológicas . 354 (1385): 903–16. doi :10.1098/rstb.1999.0442. PMC 1692594 . PMID 10382224.
^ Fayyazuddin A, Dickinson MH (octubre de 1999). "La entrada mecanosensorial convergente estructura la fase de activación de una neurona motora de dirección en la mosca azul, Calliphora". Revista de Neurofisiología . 82 (4): 1916–26. doi :10.1152/junio.1999.82.4.1916. PMID 10515981. S2CID 15194097.
^ Szczecinski, Nicolás S.; Zill, Sasha N.; Dallmann, Chris J.; Quinn, Roger D. (2020). "Modelado de las descargas sensoriales dinámicas del insecto campaniforme Sensilla". En Vouloutsi, Vasiliki; Mura, Anna; Tauber, Falk; Mota, Thomas; Prescott, Tony J.; Verschure, Paul FMJ (eds.). Sistemas Biomiméticos y Biohíbridos . Apuntes de conferencias sobre informática. vol. 12413. Cham: Editorial Internacional Springer. págs. 342–353. doi :10.1007/978-3-030-64313-3_33. ISBN978-3-030-64313-3. S2CID 230716960.
^ Orfebre, California; Szczecinski, NS; Quinn, RD (14 de septiembre de 2020). "Modelado neurodinámico de la mosca de la fruta Drosophila melanogaster". Bioinspiración y biomimética . 15 (6): 065003. doi : 10.1088/1748-3190/ab9e52. ISSN 1748-3190. PMID 32924978. S2CID 219911361.
^ ab Zyhowski, William P.; Zill, Sasha N.; Szczecinski, Nicolás S. (2023). "La retroalimentación de carga adaptativa señala sólidamente la dinámica de la fuerza en el modelo robótico del paso de Carausius morosus". Fronteras en Neurorobótica . 17 . doi : 10.3389/fnbot.2023.1125171 . ISSN 1662-5218. PMC 9908954 . PMID 36776993.