Las fibrillas (del latín fibra [1] ) son materiales biológicos estructurales que se encuentran en casi todos los organismos vivos . No deben confundirse con fibras o filamentos , las fibrillas tienden a tener diámetros que van de 10 a 100 nanómetros (mientras que las fibras son estructuras de escala micro a milimétrica y los filamentos tienen diámetros de aproximadamente 10 a 50 nanómetros). Las fibrillas no suelen encontrarse solas, sino que son partes de estructuras jerárquicas mayores que se encuentran comúnmente en los sistemas biológicos. Debido a la prevalencia de las fibrillas en los sistemas biológicos, su estudio es de gran importancia en los campos de la microbiología , la biomecánica y la ciencia de los materiales .
Las fibrillas están compuestas de biopolímeros lineales y se caracterizan por estructuras similares a varillas con altas relaciones longitud-diámetro. A menudo se organizan espontáneamente en estructuras helicoidales. En problemas de biomecánica , las fibrillas se pueden caracterizar como vigas clásicas con un área de sección transversal aproximadamente circular en la escala nanométrica. Como tal, se pueden aplicar ecuaciones simples de flexión de vigas para calcular la resistencia a la flexión de las fibrillas en condiciones de carga ultrabaja. Como la mayoría de los biopolímeros, las relaciones de tensión-deformación de las fibrillas tienden a mostrar una región característica de punta-talón antes que una región elástica lineal . [2] A diferencia de los biopolímeros, las fibrillas no se comportan como materiales homogéneos, ya que se ha demostrado que la resistencia al límite elástico varía con el volumen, lo que indica dependencias estructurales. [3] Se ha demostrado que la hidratación produce un efecto notable en las propiedades mecánicas de los materiales fibrilares. Se ha demostrado que la presencia de agua (un aldehído) disminuye la rigidez de las fibrillas de colágeno, así como aumenta su tasa de relajación de la tensión y su resistencia. [4] Desde un punto de vista biológico, el contenido de agua actúa como un mecanismo de endurecimiento de las estructuras fibrilares, permitiendo una mayor absorción de energía y mayores capacidades de deformación.
Las propiedades de fortalecimiento mecánico de las fibrillas se originan a nivel molecular. Las fuerzas distribuidas en la fibra son la carga de tracción soportada por la fibrilla y las fuerzas de corte que se sienten debido a la interacción con otras moléculas de fibrilla. La resistencia a la fractura de las moléculas de colágeno individuales está controlada por la química covalente entre moléculas. La resistencia al corte entre dos moléculas de colágeno está controlada por interacciones débiles de enlaces dispersivos y de hidrógeno y por algunos enlaces cruzados covalentes moleculares . El deslizamiento en el sistema se produce cuando estos enlaces intermoleculares enfrentan una tensión aplicada mayor que su fuerza de interacción. [5] La ruptura de enlaces intermoleculares no conduce inmediatamente a la falla, por el contrario, desempeñan un papel esencial en la disipación de energía que reduce la tensión que siente el material en general y le permite soportar la fractura. Estos enlaces, a menudo enlaces de hidrógeno e interacciones dispersivas de Van der Waals , actúan como enlaces "sacrificio", que existen con el propósito de reducir la tensión en la red. Los enlaces cruzados covalentes moleculares también juegan un papel clave en la formación de redes de fibrillas. Si bien las moléculas reticuladas pueden generar estructuras fuertes, una reticulación excesiva en las redes de biopolímeros tiene más probabilidades de fracturarse, ya que la red no puede disipar la energía, lo que genera un material fuerte pero no resistente. Esto se observa en el colágeno deshidratado o envejecido, lo que explica por qué con la edad los tejidos humanos se vuelven más frágiles [6].
Las diferencias de estructura entre fibrillas de distinto origen se determinan normalmente mediante difracción de rayos X. [2] Se puede utilizar un microscopio electrónico de barrido (SEM) para observar detalles específicos en especies de fibrillas más grandes, como las bandas características de 67 nm en el colágeno, pero a menudo no es lo suficientemente fino para determinar la estructura completa.
Los materiales naturales muestran una combinación de propiedades mecánicas normalmente contradictorias ( suavidad y tenacidad ), debido a sus estructuras jerárquicas de fibrillas en múltiples escalas de longitud. [7] Estas fibrillas a menudo están orientadas en una sola dirección, lo que genera una respuesta mecánica anisotrópica en el material biocompuesto resultante. [8] Esta es una ventaja primordial, ya que la mayoría de estos materiales resisten tensiones en una sola dirección, por lo que un mayor rendimiento y tensión de fractura en la dirección de la tensión aplicada garantiza la integridad estructural del material. Las macro, micro y nano fibrillas permiten que el material resista la fractura a través de una serie de mecanismos de resistencia a la fractura:
Estos mecanismos trabajan juntos para resistir la fractura, lo que permite que estos materiales soporten millones de ciclos de carga sin fallar, algo esencial para los seres vivos móviles. Otra ventaja mecánica de los biopolímeros es su capacidad para ser deformados, lo que resulta de la existencia de estructuras fibrilares fuertes en un material de matriz más flexible. La buena deformabilidad de las matrices interfaciales desempeña un papel clave al permitir la reorientación de los constituyentes durante la deformación. [8]
La fibrilogénesis es la expansión de fibrillas finas que es común en las fibras de colágeno del tejido conectivo . Los mecanismos definitivos de la fibrilogénesis aún son desconocidos, aunque muchas hipótesis resultantes de la investigación básica ayudan a descubrir muchos mecanismos posibles. En los primeros experimentos, el colágeno I podía destilarse de los tejidos y recombinarse en fibrillas controlando las soluciones. Estudios posteriores ayudan a comprender la composición y la estructura de los sitios de unión en los monómeros de colágeno. El colágeno se sintetiza como un precursor soluble, el procolágeno, que apoya el autoensamblaje del colágeno. Dado que las fibrillas de colágeno tienen casi 50 componentes de unión in vivo, el requisito definitivo para generar fibrilogénesis in vivo aún es críptico. [9]
Con una solución ácida o salina, el colágeno se puede extraer de los tejidos y reorganizarlo en fibrillas cambiando la temperatura o el valor del pH. [10] Los experimentos descubrieron una fuerza de atracción entre los monómeros de colágeno que ayuda a la reorganización. [11] El colágeno sirve como precursor, procolágeno, en la reacción de síntesis, que identifica la autopolimerización del colágeno.
Existen en la naturaleza más de 30 colágenos que son similares en composición química pero difieren en términos de estructura cristalina. Con diferencia, el colágeno I y II son los más abundantes. Forman fibrillas de forma proactiva in vitro, mientras que la fibronectina, las integrinas que se unen a la fibronectina y al colágeno y el colágeno V son esenciales para la formación del colágeno I y el colágeno XI para la formación del colágeno II. Por lo tanto, los mecanismos celulares desempeñan un papel clave en el proceso de autoensamblaje de proteínas.
.png/1280px-Collagen_biosynthesis_(en).png)
El colágeno es la principal proteína estructural fuera de las células en muchos tejidos conectivos de animales. [12] Como componente principal del tejido conectivo, tiene la mayor cantidad entre las proteínas de los mamíferos, ocupando entre el 25% y el 35% de todo el contenido proteico del cuerpo.
Las fibrillas de colágeno se encuentran empaquetadas en una estructura ondulada. La curva de tensión/deformación del colágeno, como el tendón, se puede subdividir en varias regiones. La región de pequeñas tensiones, la región de la "punta", corresponde a la eliminación de una ondulación macroscópica, el desenrollado, en las fibrillas de colágeno, visible en el microscopio óptico. En las tensiones más grandes, la región del "talón" y la "lineal", no hay cambios estructurales visibles.
El tropocolágeno es el componente molecular de la fibra, que consta de tres cadenas polipeptídicas zurdas (roja, verde, azul) enrolladas una alrededor de otra, formando una triple hélice dextrógira.
Los músculos se contraen y se estiran mediante el deslizamiento/agarre orientable de la miosina que interactúa con las fibras de actina . La actina consta de dos polipéptidos en una hélice y la miosina tiene una pequeña estructura en forma de corazón, un puente cruzado. Los procesos de unión y desunión de los puentes cruzados que se adhieren al filamento de actina ayudan al movimiento relativo de estos colágenos y, por lo tanto, de todo el músculo.
La elastina es una proteína fibrosa común en diversos tejidos blandos, como la piel, los vasos sanguíneos y el tejido pulmonar. Cada monómero se conecta con los demás, formando una red tridimensional, con capacidad para soportar más del 200 % de tensión antes de deformarse. [13]
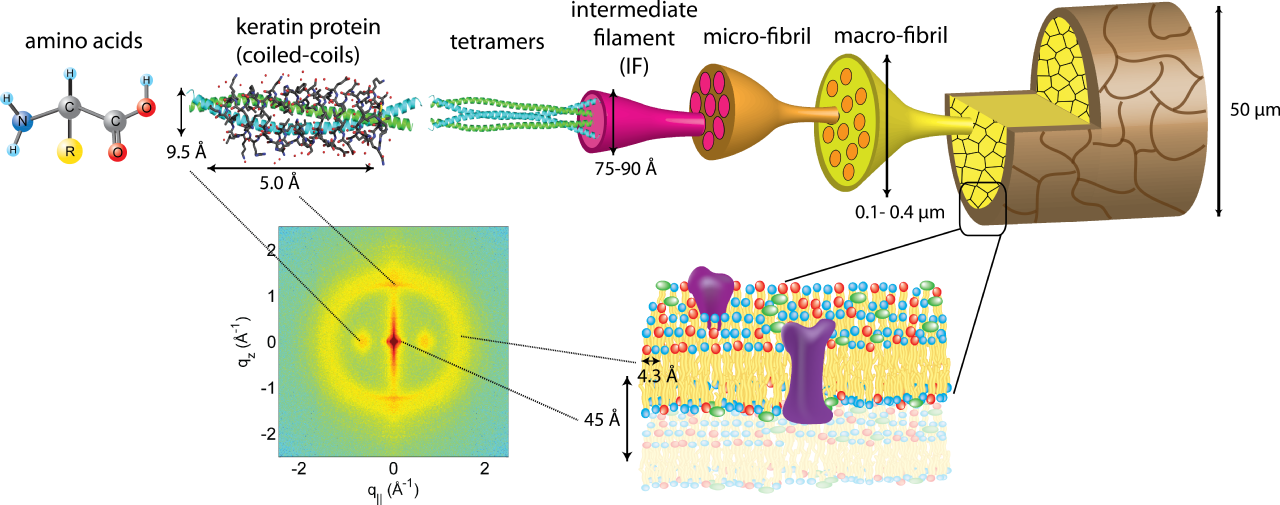
La queratina es una proteína estructural que se encuentra principalmente en el cabello, las uñas, las pezuñas, los cuernos y las púas. [14] Básicamente, la queratina está formada por cadenas polipeptídicas, que se enrollan en hélices α con enlaces cruzados de azufre o se unen en láminas β unidas por enlaces de hidrógeno. La β-queratina, que es más resistente que la conformación α, es más común en aves y reptiles.
La resilina es una proteína elastomérica de insectos que consta de estructuras de hélices α y láminas β. [15] Es una de las proteínas más resilientes de la naturaleza. Tiene una rigidez baja (~0,6 MPa) pero un alto porcentaje de recuperación de energía (~98 %), y ayuda de manera eficiente a los insectos voladores a batir sus alas o a las pulgas a saltar.
La fibrilla de seda de araña está compuesta por una estructura de láminas β cristalizadas rígidas, responsables de la resistencia, y una matriz amorfa que la rodea, lo que mejora la tenacidad y la capacidad de elongación. [16] Tiene una resistencia a la tracción y una ductilidad excepcionalmente altas, con una densidad respectivamente baja, en comparación con otras fibrillas naturales. Su característica varía de un tipo de araña a otro para diferentes utilidades.
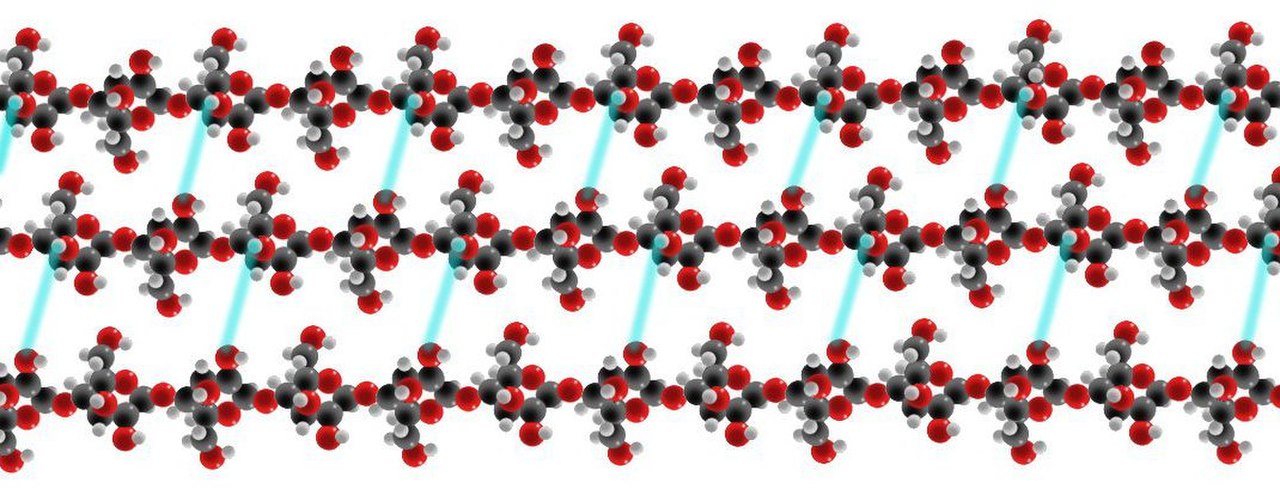
La pared celular primaria obtiene su notable resistencia a la tracción de las moléculas de celulosa, o de las largas cadenas de residuos de glucosa estabilizadas por enlaces de hidrógeno . [17] Se observa que las cadenas de celulosa se alinean en matrices paralelas superpuestas, con una polaridad similar que forma una microfibrilla de celulosa. En las plantas, estas microfibrillas de celulosa se organizan en capas, formalmente conocidas como láminas, y se estabilizan en la pared celular mediante moléculas de glicano reticuladas superficiales y largas. Las moléculas de glicano aumentan la complejidad de las redes potenciales en las que puede configurarse la celulosa vegetal. La pectina , que es un polisacárido que contiene muchas unidades de ácido galacturónico con carga negativa, es coextensiva en la pared celular primaria tanto de las microfibrillas de celulosa como de las redes de glicano complementarias. [17] Además, las microfibrillas de celulosa también contribuyen a la forma de la planta a través de la expansión celular controlada. La disposición estereoscópica de las microfibrillas en la pared celular crea sistemas de presión de turgencia que, en última instancia, conducen al crecimiento y la expansión celular. Las microfibrillas de celulosa son macromoléculas matriciales únicas, ya que son ensambladas por enzimas de celulosa sintasa ubicadas en la superficie extracelular de la membrana plasmática. [17] Se cree que la planta puede “anticipar su morfología futura controlando la orientación de las microfibrillas” mediante un mecanismo en el que las microfibrillas de celulosa se disponen sobre una matriz cortical de microtúbulos.
Se dice que al agitar una muestra determinada de amilosa se forman cristales fibrilares que se dice que se precipitan del licor madre. Estas largas fibrillas se pueden visualizar mediante microscopía electrónica y revelan estrías transversales que se asemejan a un kebab . [ cita requerida ] Las fibrillas de amilosa se clasifican según su morfología: unas con fibrillas pequeñas en forma de varilla y otras con cristales en forma de listón.
Se dice que la estructura fibrilar de la madera desempeña un papel importante tanto en la estabilidad mecánica como en la capacidad de la madera para poseer canales para transportar minerales y agua. Se informa que la madera de abeto (Picea abies), entre otras, posee fibrillas de celulosa con un diámetro normalizado de 2,5 nm. También se ha informado de un vínculo entre la edad de la madera y el ángulo espiral de las fibrillas con respecto a la dirección longitudinal. Se dice que la madera temprana tiene un ángulo de reposo constante de 4,6 ± 0,6°, mientras que la madera tardía tiene una región de transición de 4,6° a 19,8 ± 0,7°. [18] En la madera tardía, las dos regiones de ángulo espiral de las fibrillas de celulosa no son continuas, lo que significa que hay dos estructuras traqueidas independientes en los árboles "más viejos" que cumplen diferentes requisitos mecánicos. Además, las fibrillas orientadas longitudinalmente mejoran la resistencia a la tracción, mientras que la adición de fibrillas inclinadas 20°, exclusivas de las traqueidas de la madera tardía, proporciona estabilidad contra la compresión. [18]
Para imitar la fuerte adhesión, el desprendimiento fácil y las propiedades de autolimpieza de la almohadilla de los dedos de un geco, se puede crear un adhesivo a base de fibrillas. Estas características de rendimiento se derivan de la estructura jerárquica subyacente que consta de un millón de microfibrillas llamadas setas que a su vez constan de miles de millones de ramas de tamaño nanométrico llamadas espátulas .
Imitar este fenómeno implica cuatro pasos de diseño distintos: [19]
Para imitar una matriz ósea madura, se pueden utilizar fibrillas autoensambladas para alinear una matriz mineral determinada. Esto se logra utilizando una molécula autoensamblada con una cola de alquilo hidrofóbica y una cabeza de oligopéptido hidrofílica. Estas moléculas forman estructuras micelares in situ y puentes disulfuro a bajo pH, lo que conduce a la formación y cristalización de nanofibrillas poliméricas de 200 kDa. [ cita requerida ] La matriz mineral finalmente interactúa con la fibrilla sintética a través de un residuo de fosfoserina que da como resultado la nucleación y el crecimiento mineral.
{{cite journal}}: Requiere citar revista |journal=( ayuda )